 Andreas R. Hassl
Andreas R. Hassl
IV.0 Der Zecken Lebensweise
Auf dieser in den Fachbereich der Allgemeinen Zeckenkunde einführenden Internet-Seite wird ein gestraffter Überblick über die Lebensweise dieser Tiergruppe in ⇒ Zentraleuropa, speziell in Österreich dargelegt und so soll das Wissenswerteste zum Verhalten und zu den Eigenarten der Mitglieder dieser Tiergruppe allgemeinverständlich veranschaulicht werden. Unter dieser Thematik subsumiert sind Daten zu der Zecken Verhalten beim Nahrungserwerb und der Wirtssuche, ihrer jahreszeitlichen Aktivitätsphase, ihrem Partnersuchverhalten und ihrer Art des Nachkommen-in-die-Welt-Setzens.
- IV.1Die Wirtssuche
- ❍Die Wirtigkeit
- ❍Die drei Wirtsfindungsstrategien
- ❍Die Wirtsspezifität
- IV.2Der jahreszeitliche Aktivitätszyklus
- IV.3Die Nahrungsaufnahme
- ❍Der Saugakt der Schildzecken
- ❍Der Saugakt der Lederzecken
- ❍Die Nährstoffaufnahme und Verdauung
- ❍Nach dem Saugen
- IV.4Die Fortpflanzung
- ❍Die Partnerfindung und die Pheromone
- ❍Die Begattung
- ❍Partheno- und Paedogenese
- ❍Das Eierlegen
IV.1 Die Wirtssuche
Die Wirtigkeit
Die meisten Schildzecken sind relativ wirtsunspezifisch. Dies liegt vermutlich daran, dass sie im Wesentlichen nur das Hämoglobin, das Albumin, Globuline und Cholesterin zu ihrer Ernährung benötigen. Diese Substanzen sind im Blut aller Wirbeltiere enthalten. Nach wie vor ungeklärt ist die Rolle, die Symbionten im Zeckendarm bei der Verdauung des Wirbeltierblutes spielen. Diese Symbionten können in ihrer Physiologie so spezifisch sein, dass ein Parasit aus dem Stamm ⇒ Arthropoda bei Aufnahme eines anderen Blutes als desjenigen, auf das er spezialisiert ist, innerhalb weniger Stunden abstirbt. Ein Beispiel dafür sind die humanpathogenen Läuse. In solchen Fällen steht die Frage im Raum, ob eine primäre hohe Wirtsspezifität die Ausbildung einer hocheffektiven, aber störungsanfälligen Symbiontenfauna fördert, oder ob der Vorteil eines breit angelegten Verdauungssystems die Entwicklung eines Wirts-Generalisten erzwingt. Den entwicklungsgeschichtlich fortschrittlichen Weg, den zu einer hohen Wirtsspezifität bei möglichst geringer Wirtswechselfrequenz, gehen nur wenige Zecken-Arten. Solche Parasiten sind gekennzeichnet durch eine starke Reduktion ihrer nicht mehr benötigten Organe, zB der Sinnesorgane. Eine Vorstufe dazu sind Parasiten, die in ihrem Individualleben nur ein Wirtsindividuum invadieren oder in bzw an einem Wirtsindividuum lebenslang verbleiben oder im Quartier des Wirtes leben und zwar verschiedene Wirtsindividuen, aber immer nur eine Wirtstier-Art befallen. Der letztgenannte parasitische Lebensmodus ist bei Zecken besonders häufig anzutreffen. Der Grund dafür ist wohl, dass die meisten Wirbeltiere auf einen Zeckenbefall immunologisch mit der Bildung von Abwehrstoffen reagieren und nach einiger Zeit der - wohl immer nur artgleichen - Zecke das Blutsaugen durch ihre Immunabwehr weitgehend verwehren.
Dem Dilemma des Auftretens von sich gegen den Befall immunologisch wehrenden Wirten entgeht die Zecke durch:
• Einen regelmäßigen Wechsel der Wirtstier-Art bei geringstmöglicher Häufigkeit des Blutsaugens. Ist der Wirt aber geeignet, kann das Blutsaugen so lange dauern, bis der Wirt immun wird.
• Eine möglichst kurze Saugdauer, die wenig immunologisch reaktive Substanzen in den Wirt spült. Dafür muss aber dann häufiger gesaugt werden, um die für das Durchlaufen der Ontogenie benötigte Gesamtblutmenge zu erlangen.
Während Schildzecken ihren als geeignet befundenen Wirt nie, einmal oder höchstens zweimal wechseln, diese nennt man dann entsprechend ein-, zwei-, oder drei-wirtige Arten, und der Saugakt mehrere Tage bis mehrere Monate dauern kann, wechseln Lederzecken ihre Wirtsindividuen mehrfach, saugen dafür aber nur wenige Minuten bis Stunden. In vielen Schildzeckengattungen, wie zB Ixodes, gibt es einen starken Trend dazu, dass die ⇒ adulten männlichen Tiere gar nicht mehr saugen.
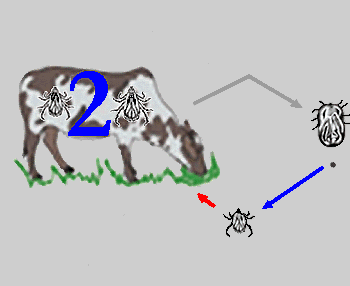
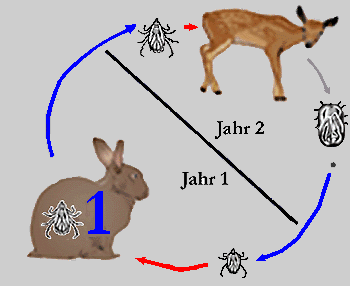
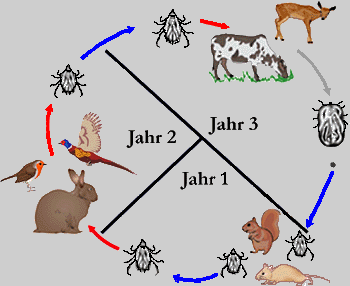
Abb. IV.1: Die Wirtigkeit von Schildzecken: Ein-, Zwei- und Dreiwirtige Zecken und deren Lebenszyklen. Blaue Pfeile repräsentieren Häutungen, rote Pfeile eine Wirtssuche zum Zweqcke der Nahrungsaufnahme. Die blauen Zahlen geben die Anzahl der Häutungen am Wirt an. In die Wirtsdarstellung gezeichnete Zeckenstadien repräsentieren Stadien, die sich am Wirt häuten. Nicht in den Wirt gezeichnete Stadien zeigen Häutungen im Freiland an. Das vollgesogene Weibchen verlässt immer den Wirt, um am Boden ihre Eier zu legen (grauer Pfeil).
© A. Hassl.
Die drei Wirtsfindungsstrategien
Dem Wechsel der Wirtsindividuen von mehrwirtigen Zecken-Arten und dem geregelten Wechsel der Wirts- , also der ⇒ telotropischen Lebensweise, stehen die für das Überleben notwendige Ausnutzung der „Jagdgründe” und entsprechend angepasste Jagdstrategien der Zecken gegenüber. Es lassen sich drei grundsätzlich unterschiedliche Strategien unterscheiden. Diese können bei verschiedenen Zecken-Arten und -gattungen, bedingt durch die unterschiedliche Anatomie, zwar abgewandelt werden, in keinem bekannten Fall werden diese Strategien jedoch vermischt. Die drei dargelegten Strategien finden sich bei den österreichischen Zecken-Arten, wobei sämtliche heimischen Lederzecken Nestschmarotzer sind. Die vierte Strategie, die eines ⇒ permanenten Ektoparasiten, kann man in Österreich nicht studieren, weil man in diesem Land bislang noch nie ein-wirtigen Schildzecken gefunden hat.
1. Der Nestschmarotzer. Die nestschmarotzende Zecke verbringt ihr ganzes Leben in oder in der unmittelbaren Umgebung der Ruhe-, Schlaf- oder Brutstätten ihrer Wirte. Die Wirtstiere kommen von sich aus zur Zecke, diese muss nur geeigneten Wirt von Unerheblichem unterscheiden können. Europaweit gesehen verhalten sich die Angehörigen der meisten Zecken-Arten so, dh sie leben vor widrigen äußeren Einflüssen weitgehend geschützt in den Nestern ihrer Wirte. Diese Lebensweise nennt man ⇒ nidikol. Leben die Zecken auch noch in höhlenartigen Strukturen, sind sie also auch noch geschützt vor Niederschlägen und den jahreszeitlichen Temperaturschwankungen, so ist ⇒ kavernikol die angebrachtere Bezeichnung, im Falle eines Lebens in Häusern oder Ställen von Haustieren ist die gängige Bezeichnung ⇒ „indoor lebend”. Diese Lebensweise scheint besonders innerhalb der Gattung Ixodes weit verbreitet zu sein. Es handelt sich erkennbar um eine sehr erfolgreiche Überlebensstrategie, die meist entweder mit einer Spezialisierung auf nur eine Wirtstier-Art einhergeht, und/oder aber Kolonie-bildende Wirte betrifft, zB die gemischtartigen Seevogelkolonien. Nidikole Tiere benötigen wenig ausgeprägte Befähigungen zum Auffinden und zum Erkennen eines geeigneten Wirtes und sie sind daher kaum zum Einsatz ausgefeilter Wirtssuchstrategien befähigt. Soweit bekannt, halten sich auch alle europäischen Lederzecken in der Nähe von den Plätzen auf, an denen ihre Wirte ruhen oder schlafen. In manchen Fällen, insbesondere im Falle von Mitgliedern einer Höhlenfauna, kann das „In-der-Nähe-Verbleiben” allerdings auch mehrere Meter einer für Zecken sehr schwierig zu überwindenden Strecke bedeuten. Solche Tiere haben dann spezielle Anpassungen, zB lange Schreitbeine und einen spinnenähnlichen Habitus. Ein Beispiel dafür ist Argas (Carios) vespertilionis.
Die zwei medizinisch bedeutsamsten Zecken-Arten Österreichs, Ixodes ricinus und Dermacentor reticulatus, sind „frei-lebend”. Sie verbringen den größten Teil ihres Lebens in der Umwelt, in oder auf den obersten Bodenschichten oder auf der bodennahen Vegetation. Im englischen Sprachraum wird das häufig als "off-host" Leben bezeichnet. Vertreter solcher Arten müssen aktiv nach einem Wirt suchen, sie sind also Jäger. Entomologen unterscheiden unter den frei-lebenden Zecken zwischen zwei grundverschiedene Jagdstrategien:
2. Der Pirschjäger. Die in Österreich vorkommenden Dermacentor-Arten, D. marginatus und D. reticulatus sind Zecken, die sich analog zu katzenartigen Raubsäugetieren verhalten. Üblicherweise verbergen sie sich in den obersten Bodenschichten. Bei Hunger und passenden Witterungsbedingungen graben sie sich aus dem Boden hervor und durchstreifen dann im Laufen ein Areal von ca 30m2; auf der Suche nach einem passenden Wirt. Sie werden daher auch „Laufzecken” bezeichnet. Über ihr Wirtserkennungssystem ist nicht viel bekannt, sie besitzen allerdings gut entwickelte Augen. Auch wird immer wieder im Zusammenhang mit den Jagdstrategien der Geruchssinn als Orientierungshilfe genannt. Vom Geruchssinn weiß man, dass Azeton, NO und CO₂ sehr attraktiv für Zecken sind, wobei Azeton und NO weniger attraktiv sind als CO₂, hingegen Isopren und NH₃ keinerlei Wirkung zeigen. In einer Studie konnte gezeigt werden, dass der Atem eines Landwirbeltieres der beste Stimulus ist. Er zog neben Amblyomma variegatum, Rhipicephalus sanguineus und Ixodes ricinus auch den Argasiden Ornithodorus moubata an. Es scheint so, dass die Evolution des Wirtssuchverhaltens von Zecken sensorische Anpassungen und Verhaltensänderungen inkludiert. Der Zweck könnte sein, Stoffwechselprodukte des Wirtes, die dieser regelmäßig ausatmet, zu erkennen und sich dann an diesen zu orientieren. cit. McMahon et Guerin [2002]. Pirschjagende Zecken haben in jedem Fall ein breites Wirtsspektrum. Zumindest in Bezug auf das Wirtsfindungsverhalten scheint es sich bei den pirschjagenden Zecken um sehr ursprüngliche, noch an räuberische Milben und deren Jagdverhalten erinnernde Formen zu handeln.

Abb. IV.2: Die Lauer- und die Ruhehaltung des Holzbocks.
3. Der Lauerjäger. Insbesondere von der Art Ixodes ricinus ist das lauernde Jagdverhalten gut erforscht und wird im Folgenden beschrieben: Zum Finden eines Wirtes klettert der Holzbock bei passender Lufttemperatur und Luftfeuchte vom Erdboden hinauf bis zur Spitze eines Grashalms oder einer anderen Pflanze. Vermutlich ist dies ein Fall von positiver ⇒ Gravitaxis. Dort können sie, gesteuert vom Instinkt, entweder in eine- ung oder in die Lauerstellung gehen. In der Ruhehaltung sind die beiden vordersten Beine in Richtung Capitulum gefalten und liegen nahe beim Körper. In der Lauerhaltung ist das vorderste Beinpaar nach oben gestreckt und wird hin und her geschwenkt. Diese Schwenkbewegungen kann man auch beim Laufen beobachten, weil die Zecke so mit ihrem Haller`schen Organ Geruchsreize aufnehmen kann und sich im Raum orientiert. Entgegen einer weitverbreiteten Meinung sitzen die lauerjagenden Zecken nicht auf den Bäumen, um sich von dort auf ihre Wirte hinunter zu stürzen, sondern sie befinden sich in der bodennahen Vegetation. Larven sind in der Regel auf Gräsern bis maximal 30 cm Höhe, Nymphen auf weniger als ein Meter hohen Kräutern, und Adulte auf Kräutern und Büschen bis zu maximal 1,5 m Höhe anzutreffen. Die Zecken sitzen meist auf der Unterseite der Blätter und an den Spitzen von Zweigen in der Nähe von Pfaden und Wildwechseln. Lauerjagende Schildzecken, die im Vergleich zu den Laufzecken relativ wirtsspezifisch sind, suchen sich Orte, die der Bauchhöhe ihres bevorzugten Wirtstieres entsprechen. Die Zecken nehmen gewöhnlich vorerst die Ruhestellung ein, bei Vibration, Geruchsreizen oder bei Änderung der Beleuchtungsstärke, besonders bei einem plötzlichen Abfall, wechseln sie sofort in die Lauerstellung. Berührt eine vorerst passende Struktur das erste Beinpaar, so lässt die Zecke mit den Hinterbeinen ihre Unterlage los und klammert sich mit den Vorderbeinen fest. Sodann beginnt sie mit der Prüfung der Struktur, auf der sie nun sitzt, auf deren Tauglichkeit als Wirt. Da dies eine Vorbereitungshandlung für den Saugakt ist, finden sich Details dazu im Kapitel: VI.3 Der Saugakt bei Schildzecken
Die Wirtsspezifität
Obgleich die Wirtsspezifität der einzelnen Zecken-Arten der entscheidende Faktor für die Begrenzung ihrer Ausbreitung, ihre lokale Verbreitung und ihre ⇒ Abundanz ist, ist der Begriff schlecht und uneinheitlich definiert. Sehr häufig erfolgt eine Vermengung und/oder sogar Synonymisierung mit dem Blutsaugen an einem Wirtstiertaxon, also der ⇒ -phagie. Meist wird der Begriff Wirtsspezifität relativ zur Spezifität des Zecken-Taxons verwendet: Moderate Wirtsspezifität zB kann bei ⇒ ornithophilen Argasiden bedeuten, dass sie einige wenige Arten von (Haus-)Geflügel als Blutspender akzeptieren, bei den meisten heimischen Ixodes-Arten hingegen, dass sie auf allen Arten von Kleinsäugetieren parasitieren, jedoch nur selten auf Großtieren, Vögeln und Reptilien gefunden werden. Vernünftig erscheinen mir folgende Präzisierungen:
Sehr strenge Wirtsspezifität oder Stenoxenie bzw homogene Polyxenie: Die Zecken-Art saugt nur an einer Wirtstier-Art Blut, kann nur dieses Blut verdauen und sich nur mit Hilfe dieses Bluts häuten. Homogene Polyxenie bedeutet, die Zecken-Art nutzt in ihrem Individualleben nur eine Wirtstier-Art, innerhalb derer allerdings mehrere Individuen. Dies ist bei zwei- oder drei-wirtigen Zecken mit nur einer Wirtstier-Art als Blutspender immer der Fall, weil jedes Stadium einen neuen Wirt sucht. Ein-wirtige Zecken, die naturgemäß immer ⇒ stenoxen sind, gibt es in Österreich nicht. In Wahrheit bedeutet Stenoxenie allerdings, die Zecke erkennt nur eine Wirtstierspezies als potentieller Blutspender. Ihre Verfahren zur Wirtserkennung sind also entweder hochspezifisch oder aber die Zecke nutzt gar keine solcher Verfahren, und ihr Verdauungsvermögen ist wenig flexibel, aber möglicherweise eben hoch effizient.
Strenge Wirtsspäzifität oder Oligoxenie: Die Zecken-Art saugt unter Freilandbedingungen an einigen wenigen Wirtstierarten Blut. Offen bleibt, ob sie alle diese Blutspender auch als Nahrungsquelle im Rahmen ihrer Ontogenie nutzen kann, oder aber ob sich eine stenoxene Zecken-Art einfach nur bei der Wirtserkennung „geirrt” hat.
Moderate Wirtsspezifität, in sehr unglücklicher Weise manchmal ebenfalls Oligoxenie genannt: Die Zecken-Art saugt an vielen verschiedenen Wirtstier-Arten Blut, bevorzugt aber Angehörige nur eines oder weniger höherer Taxa. Meist dann verwendet, wenn verschiedene Wirtstier-Arten innerhalb einer Ordnung oder einer Familie bevorzugt attackiert werden.
Geringe Wirtsspezifität oder Euryxenie: Die Zecken-Art saugt an vielen verschiedenen Wirtstier-Arten Blut. Meist dann verwendet, wenn verschiedene Klassen an Wirtstieren unterschiedslos attackiert werden.
Ein zeckenkundlich überaus faszinierendes Phänomen ist der ⇒ Telotropismus einiger Schildzecken-Arten: Die Zecken-Art ist eury- oder ⇒ polyxen, ihre Lebensstadien sind allerdings jeweils moderat wirtsspezifisch oder ⇒ oligoxen, sie nutzen sich in ihrer ⇒ Ökologie erheblich unterscheidende Wirte. Telotropismus kann auch so definiert werden: Die verschiedenen Lebensstadien einer Schildzecke parasitieren an unterschiedlichen Wirtstier-Spektren. Die einzelnen Stadien solcher Zecken werden auf unterschiedliche Weisen verschleppt, sie haben daher unterschiedliche Bedeutungen in ihrem jeweiligen Stadium-spezifischen Ökosystem.
In vielen Fällen ist eine Abtrennung der Spezifität der Zecken-Art hinsichtlich der phoretischen Wirte von den Blutspendern nicht notwendig. Denn, bei konsequenter Anwendung des Trennungsprinzips ist der Text mit sehr viel redundanter Information überladen. Deshalb wird auch hier eine derartige Trennung nicht durchgeführt, obgleich in einigen Fällen von in Österreich heimischen Zecken zumindest nur an einem Geschlecht oder an den Jungtieren der Wirte Blut gesaugt wird, während die Wirtsspezifität sich auf alle Lebensphasen der Wirtstier-Art bezieht. Das Blutsaugen am „falschen” Wirt kann für die Zecke sehr unangenehme Folgen haben: Manchmal können sie danach das aufgenommene Blut nicht vollständig verdauen, es kann den Darm verstopfen und die Zecke letztendlich daran sterben. Häufig können sie sich jedoch lediglich nicht weiter entwickeln sondern verharren im gleichen Stadium. Falls das verfehlte Stadium das Überwinterungsstadium ist, wird die Zecke beim Überwintern sterben. Gelegentlich findet man äußere Missbildungen, die auf das Blutsaugen an ⇒ Fehlwirten zurückgeführt werden.
IV.2 Der jahreszeitliche Aktivitätszyklus
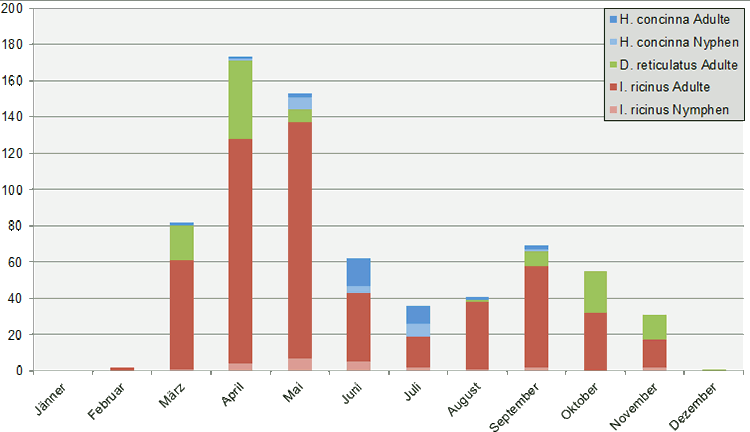
Abb. IV.3: Der jährliche Aktivitätszyklus. © A. Hassl, 2018.
Die Aktivität der Schildzecken, gemessen am Verhalten der Wirtssuche, ist in Österreich nicht gleichmäßig über das warme Halbjahr verteilt. Jede einzelne Zecken-Art zeigt im Laufe eines Jahres ein oder mehrere Aktivitäts-Maxima. Darunter werden üblicherweise Perioden verstanden, in denen die Tiere aktiv nach den Wirtsorganismen suchen und willens sind, Blut zu saugen. Die Aktivität der frei-lebenden Zecken ist hauptsächlich von der Umgebungstemperatur und der relativen Luftfeuchtigkeit abhängig und damit ein Spiegelbild der mikroklimatischen Bedingungen und der topografischen Lage des Lebensraums. Die Abbildung auf der linken Seite wurde nach Angaben von Duscher et al. [2013] zusammengestellt. Sie betrifft die Feststellung der Zahl an Hunden, die im Freiland spazieren geführt wurden, Blut saugender Schildzecken im Burgenland in Monatsperioden, sortiert nach Arten und -stadien. Man beachte, dass manche Zecken im November und Dezember aktiv sind und manche im Hochsommer eine Ruhepause einlegen.
Der temporale Aktivitätsverlauf des Holzbocks in ⇒ Zentraleuropa ist ⇒ bimodal: Es gibt ein Maximum im Frühjahr und eine zweite, kleinere Aktivitätsspitze im Herbst. Die Adulttiere und die Larven sind vorwiegend im Frühjahr aktiv, Mai bis Juni, während die Nymphen in der Regel ab April anzutreffen sind. Die zweite Aktivitätsperiode wird von alle Entwicklungsstadien von Mitte August bis in den September hervorgerufen. Diese zweigipflige Verteilung ist für die gemäßigten Breiten ⇒ ⇒ Zentraleuropas typisch. Allerdings ändert sich dies zu einer unimodalen Jahresaktivität im Falle von milden Jahresdurchschnittstemperaturen und in wärmeren Regionen Europas. Typischerweise werden unimodale ⇒ Aktivitätsmuster von Zecken vorwiegend bei Tieren beobachtet, die in geschützten ⇒ Habitaten leben, während bimodale ⇒ Aktivitätsmuster bei frei-lebenden Zecken auftreten.
Im Falle des Auftretens ungünstiger und/oder entwicklungshemmender Bedingungen können frei-lebende Zecken in einen Zustand sehr geringer metabolischer Aktivität verfallen: Der ⇒ Diapause. Dazu ziehen sich die Tiere in die ersten wenigen Zentimeter der oberflächlichen Bodenschichten zurück. Dabei kann man zwei Formen der ⇒ Diapause unterscheiden: Die „behavorial diapause” und die „morphogenetic diapause”. Mit der ersten, der Verhaltensdiapause, überbrücken nüchterne Zecken ungünstige klimatische Bedingungen, während die zweite, die ⇒ Morphogenetische Diapause, den vollgesogenen Zecken eine verzögerte Entwicklung zum nächsten Stadium ermöglicht. Korrigiert cit. Schulz [2013].
| Abkürzungen und Erläuterungen zu Tab. IV.1: | |
|---|---|
| L . . . Larven | n.d. . . . keine Angaben verfügbar |
| N . . . Nymphen | - . . . nicht anwendbar |
| A . . . Adulte | fett . . . Hauptaktivitätsmonat |
| W . . . Weibchen | |
| Tab. IV.1: Die Periodizität der Aktivität, gemessen in der Häufigkeit des Auffindens von Zecken in Österreich oder, subsidiär, in Zentraleuropa | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Zeckenart | Jänner | Februar | März | April | Mai | Juni | Juli | August | September | Oktober | November | Dezember |
| Argas persicus | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. |
| Argas polonicus | n.d. | n.d. | n.d. | L, N, A | L, N, A | L, N, A | L, N, A | L, N, A | L, N, A | L, N, A | n.d. | n.d. |
| Argas reflexus | N, A | N, A | N, A | N, A | N, A | N, A | N, A | N, A | N, A | N, A | ||
| Argas vespertilionis | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. |
| Dermacentor marginatus | A | L, N | L, N | L, N | ||||||||
| Dermacentor reticulatus | A | A | A | L | L, N | N, A | A | A | A | A | ||
| Haemaphysalis concinna | A | N, A | L, N, A | L, N, A | L, N, A | L, N | L, N | |||||
| Haemaphysalis inermis | A | A | A | L | L, N | L, N | N | A | A | A | ||
| Haemaphysalis parva | A | A | L, N | L, N | L, N | A | A | |||||
| Haemaphysalis punctata | L, N, A | L, N, A | A | A | A | A | L, N | L, N | ||||
| Hyalomma aegyptium | - | - | - | A | N, A | L, N, A | L, N, A | L, N, A | A | - | - | - |
| Hyalomma marginatum | L, N, A | L, N, A | L, N, A | L, N | L, N | |||||||
| Ixodes acuminatus | A | A | L, N, A | L, N, A | L, N, A | L, N, A | ||||||
| Ixodes apronophorus | L, N, A | L, N, A | L, N, A | L, N, A | L, N, A | L, N, A | ||||||
| Ixodes frontalis | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. |
| Ixodes inopinatus | A | N | A | N, A | N, A | N, A | ||||||
| Ixodes laguri | N, A | N, A | N, A | N, A | L, N, A | L, N, A | L, N, A | L, N, A | L, A | L, A | L, N, A | N, A |
| Ixodes persulcatus | L, N, A | L, N, A | L, N, A | |||||||||
| Ixodes ricinus | N, A | N, A | L | L | L, N, A | N, A | ||||||
| Ixodes trianguliceps | L | L | L | L, N, A | L, N, A | L, N, A | L, N, A | L, N, A | L, N, A | L, A | L | L |
| Ixodes simplex | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. |
| Ixodes vespertiliones | L, N, W | L, N, W | L, N, W | L, N, W | L, N, W | L, N, W | L, N, W | |||||
| Ixodes arboricola | L, N, W | L, N, W | L, N, W | L, N, W | L, N, W | L, N, W | L, N, W | L, N, W | L, N, W | L, N, W | L, N, W | L, N, W |
| Ixodes canisuga | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. |
| Ixodes hexagonus | N, A | N, A | N, A | L, N, A | L, N, A | L, N, A | L, N | L, N | L, N | L, N | L, N | |
| Ixodes lividus | L | L | N | N | A (?) | A (?) | ||||||
| Ixodes rugicollis | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. |
| Rhipicephalus sanguineus | L, N, A indoor | L, N, A indoor | L, N, A indoor | L, N, A indoor | L, N, A in- & outdoor | L, N, A in- & outdoor | L, N, A in- & outdoor | L, N, A in- & outdoor | L, N, A indoor | ; border-right-style:solid; ">L, N, A indoor | L, N, A indoor | L, N, A indoor |
| Rhipicephalus turanicus | - | - | - | - | - | - | - | - | - | - | - | - |
IV.3 Die Nahrungsaufnahme
Der Saugakt der Schildzecken
Findet eine Schildzecke einen ihr vorerst passend erscheinenden Wirt, so wechselt sie vom Untergrund auf seine Körperoberfläche oder -bedeckung. Danach krabbelt sie auf dem Fell, der Haut oder der Kleidung so lange herum, bis sie feststellen konnte, ob das Lebewesen auf dem sie krabbelt, ein zum Blutsaugen geeignetes Wirbeltier ist. Andernfalls lässt sie sich fallen oder gleitet herab. Falls das Tier ihr geeignet erscheint, sucht sie eine Stelle auf - en Haut, an der sie zustechen kann. Es ist nicht bekannt, welche lokalen Reize genau sie zum Stechen anregen.

Abb. IV.4: Eine festgesogene Ixodes-Zecke. © A. Hassl, 2018.
Hat eine Schildzecke eine passende Stelle auf der Haut gefunden, so strecken sie ihre hinteren Beine so, dass ihr Körper einen Winkel von 45 bis 60° zur Hautoberfläche des Wirtes bildet. Die Zecke fängt nun an, durch seitliche Bewegungen der ⇒ Chelizeren die Haut des Wirtes aufzuschneiden. Dazu ritzen die Chelizeren die Haut durch abwechselndes teleskopartiges Ausfahren oberflächlich an, um einen Ansatzpunkt zu schaffen. Anschließend werden beide Chelizeren abgewinkelt und gleichzeitig wiedereingezogen, um so das mit Widerhaken versehene ⇒ Hypostom in die Haut zu stoßen. Die Bewegung ähnelt der eines Brustschwimmers. Diese an die Funktion einer Ratsche erinnernde Bewegung der Chelizeren erlaubt es den Schildzecken, dynamisch in die weiche Hautmatrix des Wirts einzudringen und ihr starres Hypostom statisch und stabil zu verankern. cit. Richter et al. [2013]. Die Palpen spreizen sich dabei seitlich ab und schmiegen sich an die Haut des Wirtes. Manchmal löst sich die Zecke wiederum und sticht an einer benachbarten Stelle nochmals zu. Dieser Vorgang des Eindringens in die Haut des Wirts dauert circa zehn Minuten bis eine Stunde, bisweilen länger.
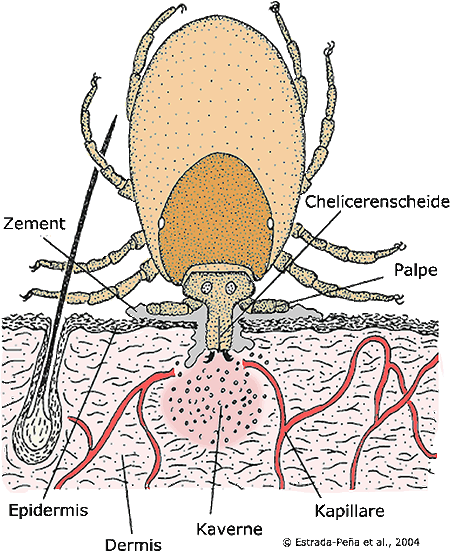
Abb. IV.5: Ein Schnitt durch die Haut während des Saugakts einer Schildzecke. ©: Estrada-Peña et al., 2004.
Das Blutsaugen einer Schildzecke: Derart in die Haut des Opfers gebohrt, sondern manche Zecken-Arten zunächst eine dickliche, weiße Flüssigkeit ab, welche schließlich ihre Mundwerkzeuge völlig umschließt. Diese verhindert eine Entzündung der Wunde und verhärtet schließlich zu einer Art Zement. In der Abbildung erkennt man die Saugkaverne, den Pool des Poolsaugers, und die Fixierung der Zecke in der Haut mittels des getrockneten Speichels, den Zement. Auf diese Weise sind die Mundwerkzeuge der Zecke derart fest im Wirt verankert, dass bei einer mechanischen Entfernung häufig das ⇒ Capitulum abreißt. Später wird diese Flüssigkeit auch direkt in die Wunde abgesondert, um die Festigkeit des Halts zu vergrößern und die Auswirkungen der Verletzung zu vermindern. Einmal auf diese Weise fest in der Haut verankert, schneidet sich die Zecke durch ruckartige Bewegungen ihrer Chelizeren noch tiefer in die Haut. Die Mundwerkzeuge stecken tief in der Haut. Nun wird eine klare Flüssigkeit (Speichel) abgesondert, welche die Gerinnung des Blutes verhindert und die Gewebe des Wirtes lysiert (auflöst). Dadurch entsteht eine Wundhöhle, in der sich Blut, interstitielle Flüssigkeit, Zelltrümmer und Makrophagen ansammeln. Zur Ausbildung oder Vergrößerung der Saugkaverne injizieren Zecken über die Speicheldrüse spezielle Faktoren, wie Blutgerinnungshemmer (Glyko- und Mucoproteine als Thrombokinase-Inhibitoren), Cytolysine (wie Esterasen, Phosphatasen, Apyrase, Bradykinin-inaktivierendes Enzym und andere), vasoaktive Mediatoren (wie Prostaglandine) und Anti-Histaminika. Dadurch wird die Gefäßpermeabilität erhöht und der Blutfluss in die Saugkaverne aufrechterhalten. cit. Wunderlich et al. [1996]. Während des Saugakts nimmt die Zecke nun abwechselnd dieses Gemisch auf und gibt Speichel in die Wundhöhle ab, sie pumpt also Flüssigkeit hin und her. Die Aufnahme dauert 2 bis 30 - die Speichelabgabe ein bis zwei Sekunden. Dazwischen liegen inaktive Phasen von einigen Sekunden bis Minuten Dauer. Mit dem Speichel wird von der Schildzecke auch das bei der Blutaufnahme aufgenommene und sofort abgetrennte, überschüssige Wasser wieder in den Wirt gepumpt. Diese Art des Blutsaugens hat erhebliche Auswirkungen auf die Effizienz der Übertragung von jenen Krankheitskeimen, die mit dem Saugakt übertragen werden. Zecken sind verglichen mit Stechmücken ein ineffizienter ⇒ Vektor!
Der Saugakt der Lederzecken
Aufgrund der Tatsache, dass das ⇒ Capitulum bei den Lederzecken unter dem Körper verborgen liegt und deshalb eine direkte Beobachtung des Saugakts schwierig ist, ist das Wissen über die genaue Weise der Nahrungsaufnahme dieser Tiere gering. Das Einführen der Mundwerkzeuge dürfte ungefähr wie bei den Schildzecken verlaufen: Die ⇒ Chelizeren schneiden eine Öffnung in die Haut, in die das ⇒ Hypostom durch Vor- und Zurückbewegen des Körpers eingeführt wird. Lederzecken kleben sich allerdings nicht mit einer Art Zement an den Wirt. Sie haben einen halbtransparenten Speichel, der in unregelmäßigen Abständen in die Wunde entlassen wird. Dessen Wirkungen sind nicht genau bekannt. Das bei der Blutaufnahme aufgenommene und sofort abgetrennte, überschüssige Wasser wird mittels der Coxaldrüsen ausgeschieden, es gelangt also nicht mehr in den Wirt zurück.
Nahrungsaufnahme und Verdauung
Das aufgenommene Blut wird sowohl bei den Schild- wie den Lederzeck- ickt. Die Konzentration der Blutinhaltstoffe kann dabei uU auf das Zwanzigfache ansteigen. Eine isotonische Salzlösung wird zu diesem Zweck dem aufgesogenen Blut entnommen und zusammen mit dem Speichel wieder in die Wunde abgegeben. Im Falle der Schildzecken werden das Hämoglobin und die meisten anderen Proteine fast sofort von den Zellen des Mitteldarms aufgenommen, welche dadurch ihre Größe vervielfachen. Bei den Lederzecken dienen die hinteren Caeca als Hauptspeicherorgane für das eingedickte Blut. Dieses kann dort mehrere Monate verbleiben. Die Zelltypen, die an der Verdauung beteiligt sind, sind bekannt, der genaue Mechanismus der Verdauungsvorgänge ist aber zu komplex, um in diesem Rahmen dargelegt zu werden.
Nach dem Saugen
Vollgesogene Zecken lassen sich meist von ihrem Wirt fallen und verhalten sich, ebenso wie auch gerade gehäutete Zecken, stark photophobisch, dh sie flüchten vor dem Licht in den Boden, insbesondere meiden sie jede direkte Bestrahlung durch Sonnenlicht. Adulte, begattete Weibchen produzieren nun ihre Eier, die häufig in den obersten Bodenschichten abgelegt werden.
IV.4 Fortpflanzung
Partnerfindung/Pheromone
Die Partnerfindung bei den Zecken scheint zumindest zu einem guten Teil durch Geruchsstoffe (Pheromone) bewerkstelligt zu werden. Man hat bei den Zecken verschiedene Arten von Pheromonen gefunden: Es gibt Zeckenspezies, deren Angehörige Versammlungs-Pheromone (assembly pheromone) absondern können. Dies sind wenig flüchtige Stoffe, die auf die Oberflächen jener Substrate abgesondert werden, auf denen die Zecken verweilen. Trifft ein Artangehöriger auf solch einen Stoff, hört er auf, sich zu bewegen. Auf diese Weise bilden sich Ansammlungen von Zecken, im Falle der Lederzecken finden sich diese in Fels- und Mauerspalten und Vogelnestern. Vermutlich führt dieses Verhalten zur Versammlung der Tiere an für die Fortpflanzung günstigen Orten. Ein anderes geschlechtsunspezifisches Pheromon wird von einigen Schildzeckenmännchen nach einigen Tagen des Blutsaugens abgegeben. Es wirkt anziehend auf Weibchen, Männchen und Nymphen und veranlasst sie, sich in der Nähe festzusaugen. Dieses Pheromon (aggregation-attachment-pheromone) spielt nur bei Arten eine Rolle, bei der die Männchen saugen, insbesondere Arten der Gattung Amblyomma. Geschlechtsspezifische Pheromone werden bei den Schildzecken ausschließlich an gerade saugenden Weibchen abgegeben und wirken ausschließlich auf saugende oder vollgesogene Männchen. Die Männchen lösen sich in diesem Fall von ihrem Wirt und machen sich auf den Weg zum Weibchen. Die Reichweite solcher Pheromone ist nicht groß (einige Zentimeter), aber auf einem Wirt herumwandernde Männchen (und Weibchen) bevorzugen Plätze auf ihrem Wirtstier, die nach solchen Pheromonen riechen, so dass sich an diesen Stellen Ansammlungen von saugenden und vollgesaugten geschlechtsreifen Tieren bilden. Bei den Lederzecken sind solche Attraktionspheromone bisher nicht beobachtet worden. Haben sich Männchen und Weibchen gefunden, so werden sowohl bei den Schildzecken wie den Lederzecken Kontakt-Pheromone abgegeben, die die eigentliche Begattung stimulieren: Zunächst testet das Männchen anhand von Pheromonen auf dem Rücken des Weibchen, ob dieses bereits lange genug Blut gesaugt hat, um fruchtbar zu sein. Ist dies der Fall, so wird die Paarung vollzogen. Diese Pheromone werden von den Schildzecken wahrscheinlich mit dem Haller`schen Organ wahrgenommen, da im Rahmen der Partnerfindung schwenkenden Bewegungen des ersten Beinpaares ausgeführt werden. Bei den Lederzecken erfüllen chemorezeptive Härchen an den Palpen denselben Zweck.
Die Paarung

Abb. IV.6: Ein Holzbock-Paar bei der Paarung.
Über das Verhalten der beiden Partner vor und bei der Begattung ist wenig Präzises bekannt, die Beschreibung von Woolley et Tyler [1988] ist nicht leicht verständlich und lässt sich nicht ohne Ungenauigkeiten in einfache Worte kleiden. In Grzimeks Tierleben [1980] findet sich hingegen eine einfache Beschreibung des Paarungsaktes von Ixodes ricinus: Zu Begattung nähern sich die Männchen den am Wirt festgesogenen Weibchen und kriecht Bauch an Bauch unter deren Leib. Dann wird das ⇒ Hypostom in die weibliche Geschlechtsöffnung eingeführt und diese erweitert. Sodann dreht sich das Männchen um seine Querachse (männliches Capitulum zu weiblichem Hinterende) und setzt eine ⇒ Spermatophore auf der weiblichen Kutikula ab. Es nimmt diese Spermatophore mit dem Hypostom und den Pedipalpen oder aber nur mit einer seiner beiden ⇒ Chelizeren auf und führt sie in den Genitaltrakt des Weibchens ein. Diese Art der Übertragung des Samens nennt man Tacospermie. Die Spermatophoren von Zecken haben keine auffälligen Besonderheiten: Auf dem auf dem Untergrund sitzenden Schild steht ein Stiel und auf diesem sitzen zwei Samenkapseln. Die Befruchtung der Eier findet im Körper des Weibchens statt, nachdem dieses die Spermatophore in sich aufgenommen hat.
Partheno- und Paedogenese bei Zecken
Die Parthenogenese ist eine Form der geschlechtlichen (!) Fortpflanzung, bei der sich aus unbefruchteten Eizellen lebensfähige und sich weiter vermehrende Adulttiere entwickeln. Eine geschlechtliche Fortpflanzung liegt dann vor, wenn die Ausgangszelle des Organismus der nächsten Generation eine „echte” Eizelle ist, die aus einer Meiose hervorgegangen ist. Parthenogenese schränkt die Durchmischung des genetischen Materials der Art im Genpool ein. Im Tierreich findet man, vereinfacht dargestellt, zwei Arten von Parthenogen- eine, die mit, und die andere, die ohne einer Kopulation mit einem Männchen stattfindet. Erstere ist adspektorisch schwierig festzustellen und ihr Vorkommen bei Zecken daher nicht gänzlich auszuschließen. Vermutlich kommt aber bei den Zecken eine Parthenogenese nur selten vor: Manchmal und regellos entwickeln sich unbefruchtete Eier von einigen Zecken-Arten zu Weibchen, gelegentlich aber auch zu Weibchen und Männchen (Thelytokie bzw. Deuterotokie). Bekannte parthenogenetische Arten sind Amblyomma cordiferum und Amblyomma rotundatum, beides an ⇒ Herpetotaxa saugende Arten. cit. Kolonin [2004]. Für das Vorliegen einer Heterogonie, das ist ein planmäßiger Wechsel zwischen einer zweigeschlechtlichen Generation und einer oder mehrerer eingeschlechtlicher Generationen, gibt es bislang keine Beweise. Lediglich von einigen Zecken-Arten, zB von Nuttalliella namaqua, sind von den Adulten bislang nur Weibchen bekannt geworden. In diesen Fällen könnte es sich aber eher um ein Problem des Suchens am falschen Ort und zur falschen Zeit handeln, denn um eine parthenogenetische Art. Die normale Anzahl an Chromosomen (2n) liegt bei Zecken zwischen 16 und 32, nicht bekannt ist die Chromosomenzahl von parthenogenetischen Tieren.
Die Paedogenese ist eine spezielle Form der Parthenogenese. Die Fortpflanzung ist bei paedogenetischen Organismen in ein Stadium der Juvenilentwicklung vorverlegt, zB bei einigen Insekten in ein Larven- oder Puppenstadium. Bei der Paedogenese handelt es sich um Thelytokie, aus unbefruchteten Eiern entstehen immer nur Weibchen. Paedogenese wurde bei Milben und damit auch bei Zecken bislang noch nie beobachtet.
Das Eierlegen
Die Zahl der insgesamt abgelegten Eier ist bei den verschiedenen Zeckengattungen unterschiedlich. Lederzeckenweibchen legen mehrmals in ihrem Leben Eier ab, Schildzeckenweibchen grundsätzlich nur ein einziges Mal. Die Menge an Eimasse ist von der Reichlichkeit der vorherigen Blutmahlzeit abhängig, die Befruchtungsquote von der Qualität des Samens:
- Argas sp. . . . oftmals je 20 bis 100 Stück
- Nuttalliella namaqua. . . . mindestens zweimal je 80 bis 150 Stück. cit. Latif et al. [2012].
- Ixodes sp. . . . 100 bis 1 000 Stück
- Haemaphysalis sp. . . . 100 bis 5 000 Stück
- Dermacentor sp. . . . 100 bis 1 000 Stück
- Rhipicephalus sp. . . 1 000 bis 5 000 Stück
- Hyalomma sp. . . 1 500 bis 15 000 Stück
- Amblyomma sp. . . 1 500 bis 20 000 Stück
Die Eier sind eine leichte Beute für Raubmilben und andere bodenlebende Fleischfresser und ein wunderbares Substrat für zersetzende Pilze und andere Destruenten. Ein unbestätigtes Gerücht besagt, dass der Inhalt von Schildzeckeneiern extrem ⇒ toxisch auf Milben und andere Mikroorganismen wirken soll und so das Ei vom Gefressenwerden geschützt ist.