 Andreas R. Hassl
Andreas R. Hassl
III.0 Der Körperbau von Zecken
Auf dieser in den Fachbereich der Allgemeinen Zeckenkunde einführenden Internet-Seite wird ein gestraffter Überblick über das Wissenswerteste zum Körperbau dieser Tiergruppe dargelegt und so soll das Wissenswerteste zur eindeutigen Erkennung von Mitgliedern dieser Tiergruppe allgemeinverständlich veranschaulicht werden. Unter dieser Thematik subsumiert sind Daten zu der Zecken Körpergliederung, ihrer Körperhülle und deren Anhänge, den Beinen, den Sinnes- und den inneren Organen und ihren Funktionen.
Inhaltsverzeichnis:
- III.1Die Körpergliederung
- III.2Das Gnathosoma
- ❍Das Capitulum
- ❍Die Chelizeren
- ❍Die Palpen
- ❍Das Hypostom
- ❍Die Areae porosae
- III.3Das Integument und die Borstenhaare
- ❍Der Aufbau des Integuments
- ❍Die Beschilderung von Zecken
- ❍Die Borstenhaare (Setae)
- ❍Die Körperfurchen
- III.4Die Beine
- ❍Das Coxenexterieur
- III.5Die viszerale Anatomie
- III.6Die Verdauungs- und Ausscheidungsorgane
- ❍Der Darmtrakt
- ❍Die Speicheldrüsen und -gänge
- ❍Die Malpighischen Gefäße
- III.7Das Blutgefäßsystem
- III.8Das Atmungssystem
- III.9Das Zentralnervensystem
- III.10Die Sinnesorgane
- ❍Die Sinnesborsten
- ❍Das Haller'sche Organ
- ❍Die Augen
- III.11Das Geschlecht
- ❍Die Geschlechtsbestimmung
- ❍Die sekundären Geschlechtsunterschiede
- ❍Die weiblichen Geschlechtsorgane
- ❍Das Gené'sche Organ
- ❍Die männlichen Geschlechtsorgane
III.1 Die Körpergliederung
In der besonders hoch entwickelten Arthropodengruppe Milben gliedert sich der Körper in den meisten Fällen in zwei, deutlich voneinander abgrenzbare und beweglich voneinander abgesetzte Körperabschnitte: Den vorderen Abschnitt um den Mund herum nennt man im Falle der Milben ⇒ Gnathosoma. Dieses Gnathosoma trägt allerdings im Falle der Zecken aus historischen Gründen den Namen ⇒ Capitulum. Es ist ein adspektorisch gut erkennbarer Körperteil, der den Mundbereich sowie die dazugehörigen Mundwerkzeuge umfasst. Es ist eine abgeleitete Struktur, die nicht der stammesgeschichtlich hergebrachten Körpergliederung eines Gliederfüßers entspricht und innerhalb der Kieferklauenträger (Chelicerata) nur in der Gruppe der Milben ausgebildet ist. Es stellt somit eine ⇒ Autapomorphie dieser Gruppe dar. Dieser Körperabschnitt entspricht genau dem, was häufig landläufig „Zeckenkopf” genannt wird.
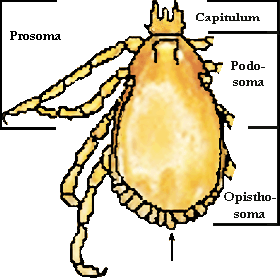
Abb. 1: Die Körpergliederung einer Schildzecke und die Festone (Pfeil).
Der übrige Körper heißt ⇒ Idiosoma. Dessen vorderer Abschnitt, dort wo die Beine ansetzen, wird von den Entomologen Podosoma genannt. Er geht im Falle der rezent existierenden Zecken ohne erkennbare Grenze in den hinteren, Bein-losen Abschnitt, das Opisthosoma, über. Diese etwas verwirrende Benennung beruht auf der Berücksichtigung des Wissens über die Stammesgeschichte der Arthropoden: Das ⇒ Capitulum und das Podosoma zusammen formen den vorderen, ursprünglich sechs Beinpaare tragenden Körperabschnitt eines fiktiven Ur-Arthropoden, der Prosoma genannt wird. Das Opisthosoma der rezenten Zecken ist hingegen in der uralten, angenommenen Gliederung des Ur-Arthropoden der beinlose, hintere Körperabschnitt. Heute lässt sich an den Zecken diese altertümliche Körpergliederung äußerlich nur mehr im Zuge einer Häutung feststellen, bei der eine temporäre transversale ⇒ Körperfurche genau an der Grenze der Abschnitte Pro- und Opisthosoma auftritt. Im Körperinneren kann die stammesgeschichtlich alte Gliederung an den dorsoventral verlaufenden Muskelsträngen nachvollzogen werden.
Im Falle einiger Schildzeckengattungen kann man möglicherweise zudem noch eine Andeutungen einer ursprünglichen Körpersegmentierung erkennen: Ihr hinterer Körperrand ist mittels Kerben in neun bis elf Abschnitte unterteilt, der Einzelabschnitt heißt ⇒ Feston. Manchmal ist die Festonreihe noch in einen größeren innersten Feston, auch Schwanzanhang oder Parma genannt, und acht bis zehn kleinere, äußere Festone gegliedert. Diese Festone sind auf dem Schema eines Rhipicephalus-Männchens linker Hand gut zu erkennen.
Im folgenden Abschnitt wird detaillierter auf die Besonderheiten der Morphologie der Schild- und der Lederzecken eingegangen, wobei als Basis das Opus von Woolley et Tyler [1988] herangezogen wird. Die beiden unten stehenden Bilder zeigen im linken Bild die Bauchansicht einer idealisierten erwachsenen Schildzecke und die einer adulten Lederzecke der Gattung Argas im rechten. Man beachte die Lage der Mundwerkzeuge als ein die rezent existierenden Familien charakterisierendes Merkmal, die bei den Lederzecken am lebenden, postlarvalen Tier unter dem Körper verborgen liegen, während sie bei den Schildzecken vor dem Körper liegen und damit gut sichtbar sind.

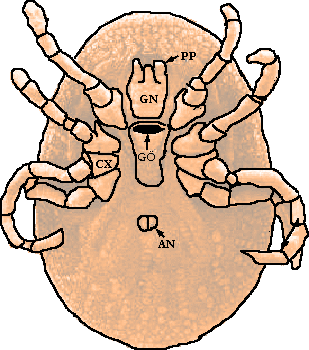
Abb. 2: Bauchseiten einer adulten Schild- und einer Lederzecke .
Abkürzungslegende: AN: Anus; CH: Chelizeren; CX: Coxa, dies ist einer der acht Beinansätze; GN: Gnathosoma (= Capitulum); H: Hypostom; GÖ: Genitalöffnung; KL: Klaue (besteht aus zwei „Krallen”); PP: Palpen; STI: Stigmen = Tracheenöffnungen (REM: Diese sind im Falle der Lederzecken auf der Bauchseite nicht sichtbar); TA: Tarsus.
III.2 Das Gnathosoma
Das Capitulum
 Das Capitulum oder Gnathosoma = Rostrum ist aus den Mundwerkzeugen und dem Kopfende einer an der Basis stehenden „Urmilbe” entstanden. Es ist ein wesentliches Merkmal für die Taxonomie und ein charakteristisches Merkmal für Zecken. Es sind die Mundwerkzeuge und nicht - wie fälschlich immer wieder geäußert wird - der „Kopf” der Tiere. Denn: Bei keinem Arthropoden liegt der Hauptteil eines Zentralnervensystems in diesem Körperteil und bei keiner Zecken-Art sitzen die Augen auf dem Capitulum. Dort gibt es ausschließlich Sinnesorgane für jene Sinne, die wir als Berührung und Geschmack bezeichnen. Das Capitulum ist der Teil des Körpers, in dem die Nahrung für die Verdauungsorgane aufbereitet wird. Selbst im Embryonalstadium kann die Segmentierung nicht mehr erkannt werden, weshalb eine exakte Homologisierung der Teile unmöglich ist. Bei den Chelicerata wurde das erste Gliedmaßenpaar zu den Mundwerkzeugen, den ⇒ Chelizeren. Diese, und nur diese, sind in dieser Tiergruppe in der Lage, Beute durch Zerzupfen zu zerkleinern. Den Chelizeren folgen nach ⇒ kaudal weitere stabförmige Extremitätenpaare, deren Coxen durch Ladenbildung zwar am Saugen beteiligt werden, jedoch niemals kauen.
Das Capitulum oder Gnathosoma = Rostrum ist aus den Mundwerkzeugen und dem Kopfende einer an der Basis stehenden „Urmilbe” entstanden. Es ist ein wesentliches Merkmal für die Taxonomie und ein charakteristisches Merkmal für Zecken. Es sind die Mundwerkzeuge und nicht - wie fälschlich immer wieder geäußert wird - der „Kopf” der Tiere. Denn: Bei keinem Arthropoden liegt der Hauptteil eines Zentralnervensystems in diesem Körperteil und bei keiner Zecken-Art sitzen die Augen auf dem Capitulum. Dort gibt es ausschließlich Sinnesorgane für jene Sinne, die wir als Berührung und Geschmack bezeichnen. Das Capitulum ist der Teil des Körpers, in dem die Nahrung für die Verdauungsorgane aufbereitet wird. Selbst im Embryonalstadium kann die Segmentierung nicht mehr erkannt werden, weshalb eine exakte Homologisierung der Teile unmöglich ist. Bei den Chelicerata wurde das erste Gliedmaßenpaar zu den Mundwerkzeugen, den ⇒ Chelizeren. Diese, und nur diese, sind in dieser Tiergruppe in der Lage, Beute durch Zerzupfen zu zerkleinern. Den Chelizeren folgen nach ⇒ kaudal weitere stabförmige Extremitätenpaare, deren Coxen durch Ladenbildung zwar am Saugen beteiligt werden, jedoch niemals kauen.
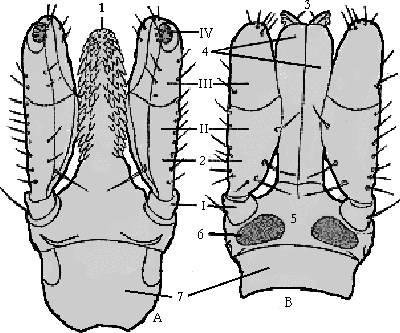
Abb. 4 & 5: Die Bilder zeigen die Capitula einer Schildzecke . Das Bild A zeigt ein Schema der ⇒ Ventral-, B eines der ⇒ Dorsalseite eines vollständigen, frei präparierten Capitulums . Das Bild zur rechten Hand ist eine Rasterelektronenmikroskop-Aufnahme. Die Nummern-Kennzeichnung der Teile ist in beiden Bildern gleich: Römisch I bis IV bezeichnen die Glieder der Palpen (2). Der basale Teil des Gnathosomas, die röhrenförmige ⇒ Basis capituli (5), ist aus der Verschmelzung mehrerer Teile entstanden. Bei den Lederzecken besteht die ⇒ Basis capituli aus einem gefalteten Kragen, der beim Saugen ausgedehnt werden kann. Bei den Schildzecken besteht sie aus den ⇒ Coxen der nun seitlich von ihren Coxen gelegenen Palpen. Der dorsale Teil des Capitulums von Schildzecken ist sklerotisiert und bildet ein Dach, das Tectum capituli (5), welches an den Seiten mit den Palpenbasen verwachsen ist. Die ⇒ Areae porosae (6) sind Drüsenareale, die nur die geschlechtsreifen Weibchen besitzen. Unterhalb der Kutikula und daher am lebenden Tier nicht sichtbar, liegen die zu einem Ring verschmolzenen maxillaren Subpalpuscoxen, Subcollare genannt (7). Die beiden ⇒ Chelizeren (3) sind messerartig geformt und mit Widerhaken besetzt. Das Hypostom (1) ist im Falle der Zecken zungenförmig ausgebildet. Die Mundöffnung liegt ⇒ median unterhalb der Chelizeren und oberhalb des Hypostoms. Sie ist sowohl in der Dorsal- als auch in der Ventralansicht am lebendigen Tier nicht sichtbar. |

|
Die Chelizeren
Die Chelizeren werden von den Zecken zum Aufschneiden der Haut des Wirtes genutzt. Die Form der Zecken-Chelizeren weicht von der der anderen Milben und auch der anderer Arachniden ab. Typischerweise bestehen sie aus drei Segmenten. Am distalen Ende der Chelizeren sitzen ein festes und ein bewegliches fingerförmiges Glied, die in der Gruppe der Milben häufig zu einer Schere umgebildet werden. Die Chelizeren der Zecken sind hochentwickelte und spezialisierte Organe, die der Penetration der Haut des Wirtes dienen. Sie bestehen entweder aus nur zwei Segmenten mit zwei beweglichen Fingern als Neubildungen oder aber aus den ursprünglichen drei Segmenten, wobei dann aber nur einer der beiden beweglichen Finger als „echtes” Endglied aufzufassen ist.

Abb. 6: Die beiden Chelizeren einer Ixodes ricinus-Larve und das darunter liegende Hypostom. ©: unbekannt.
Das ⇒ proximale Ende des ersten Segments, der Teil, welches im Körper gelagert ist, ist verbreitert und blattförmig. An ihn setzen die Muskeln an, mit denen die Chelizerenfinger bewegt werden. Die beiden Finger an der Spitze der Chelizere sind mehr oder weniger dreieckig im Umriss und lassen sich sowohl nach distal als auch seitlich bewegen. Sie tragen an den Vorder- und Seitenkanten scharfe Schneiden zum Aufschneiden der Haut. An der Basis dieser Finger setzen die Muskeln an, die zum proximalen Teil der Chelizere führen. Mit den Chelizeren kann die Haut des Wirts aufgeschnitten, genauer aufgesägt werden. Die Chelizeren werden dazu vor- und rückwärts bewegt und so eine Grube in die Haut geschnitten. Ist diese tief genug um nicht aufzureißen, klappen die Chelizeren nach seitlich außen und die Zecke rammt mittels der Hebelwirkung ihr Hypostom in die Wunde. cit. Richter et al. [2013].
Die Palpen
Die ⇒ Palpen dienen den Spinnentieren einerseits als Tastorgane und andererseits dazu, die Nahrung zur Mundöffnung zu führen. Zudem werden sie auch zur Reinigung der ⇒ Chelizeren genutzt. Bei den Zecken ist vermutlich nur die erste Funktion übrig geblieben. Milben-Palpen wurden aus den Telopoditen, den Schreitästen des ursprünglichen Spaltbeins von Arthropoden, des zweiten Extremitätenpaars gebildet. Zecken-Palpen bestehen meist aus vier Segmenten. Bei den Lederzecken sind diese Segmente ungefähr gleich lang und ohne besondere Modifikationen. Bei den Schildzecken dagegen können die einzelnen Segmente sehr ungleich groß sein. Das vorderste Segment, der ⇒ Tarsus, ist sehr klein und liegt in einer kleinen Grube des dahinterliegenden dritten Segments verborgen. Die Form der Palpen ist derart modifiziert, dass die sie häufig das Hypostom und die Chelizeren umschließen und schützen, wenn sie entlang der Längsachse des Tieres zusammen geklappt werden. Die Palpen der Zecken sind vermutlich sowohl mit Tast- als auch mit Chemorezeption ausgestattet.
Das Hypostom

Abb. 7: Die Oberseite des Hypostoms von Ixodes canisuga mit der gut erkennbaren Blutrinne. ©: unbekannt.
Das ⇒ Hypostom dient einerseits zur Verankerung der Zecke im Gewebe des Wirtes und andererseits zur Ableitung des Blutes. Es kann in einer Analogie mit der hohlen Hand eines Menschen beim Wasserschöpfen verglichen werden. Es dient - ähnlich einer Rinne an den Obstpressen - der Aufnahme und dem Transport des Blutes von der Blutkaverne bis zur Mundöffnung. Gebildet wird das Hypostom aus den ⇒ median angeordneten Verbreiterungen der ⇒ Coxen des zweiten Extremitätenpaares, der Palpen. Es ist möglich, dass sich an der Bildung des Hypostoms auch der Rest einer Unterlippe beteiligt. Diese Teile bilden zusammen einen Boden vor der Mundöffnung, der sekundär dann nach distal verlängert und in der Form umgebildet wurde. Es ist vor allem auf der Unterseite mit Widerhaken versehen, auf der in vivo nie sichtbaren Oberseite befindet sich die Rinne zur Ableitung des Blutes. Es wird beim Stichakt tief in die Haut des Wirtes versenkt und mit den Widerhaken dort verankert. Es ist daher art- und stadienspezifisch so geformt, dass es sich an die anatomischen Gegebenheiten des Wirtstieres anpasst. Insbesondere muss die Länge des Hypostoms die Dicke der Epidermis des Wirtstieres übertreffen. Die menschliche Epidermis ist etwa 0,1 mm dick, das Hypostom eines erwachsenen Holzbocks ist etwa 0,5 mm lang. Die Form und Größe des Hypostoms sind wichtige ⇒ Bestimmungsmerkmale. Weil es ein Merkmal ist, das in allen, auch den ausgestorbenen Zeckenfamilien gefunden wird, soll es ein Indiz für ein ursprüngliches Blutsaugen in diesem Taxons sein. Arten mit schnellsaugenden Vertretern zeigen eine klare Tendenz zur Reduktion der Zähne am Hypostom, nicht-saugende Stadien besitzen manchmal gar kein Hypostom (mehr).
Die Areae porosae
Auf der Oberseite der ⇒ Basis capituli, direkt hinter dem Ansatz der Palpen liegen die ⇒ Areae porosae. Diese Areale gibt es nur bei den Weibchen von Schildzecken, sie kennzeichnen diese auch. Die Areae porosae sind dicke Chitinplatten, die von Poren durchzogen sind. Bei manchen Arten sind diese Flächen unter die Ebene der ⇒ Basis capituli eingesenkt. Die Poren queren die Chitinplatte nicht vollständig. Sie enthalten ein Bündel von Borstenhaaren (⇒ Setae). Die genaue Funktion der Areae porosae ist nicht bekannt. Sie könnten ein Sinnesorgan sein oder, wahrscheinlicher, sie sind ein Areal mit Drüsenausfuhrgängen. Falls es sich um Drüsenausfuhrgänge handelt, dann haben diese Drüsen eine Funktion im Rahmen des Eierlegens. Manchmal wird behauptet, dass diese Drüsen Antioxidantien mit unbekannter Funktion absondern.
III.3 Das Integument und die Borstenhaare
Der Aufbau des Integuments
Milben sind von einem Integument, der Körperhülle, einem analogen Organ zur Haut eines Säugetiers, umhüllt. Das Integument ist ein typisches Merkmal aller Gliedertiere (⇒ Arthropoda). Eine aus nur einer Zellschicht bestehende Hypodermis (auch Epidermis) sondert Chitin und Proteine ab, aus denen sich das Exoskelett zusammensetzt. Dieses gibt dem Milbenkörper seine Form und dient den Muskeln als Ansatzareal.
Alles, was außerhalb der Hypodermis liegt, heißt Cuticula. Diese besteht aus Sklerotin, einem Komplex aus verfestigten Proteinen und Chitin. Die Cuticula gliedert sich in eine innen liegende Procuticula und eine außen liegende Epicuticula. Die Procuticula besteht aus vielen, in Schichten angeordneten Chitinlamellen. Man unterscheidet wiederum zwei größere Lagen: Die innen liegende Endocuticula und die außen liegende Exocuticula. Die Epicuticula besteht aus einer wachsartigen Außenhülle, dem Tectostracum und einer meist farbigen Innenschicht, dem Epiostracum, welches aus Chitin besteht. Von außen nach innen gereiht kann man also folgende Schichten feststellen:
- Epicuticula
- Tectostracum
- Epiostracum
- Procuticula
- Exocuticula (früher: Ectostracum)
- Endocuticula (früher: Hypostracum)
- Hypodermis (Epidermis)
Durch das ganze Integument ziehen sich Porenkanäle. Im ⇒ Taxon Schildzecken ⇒ hypertrophieren die Hypodermiszellen während des Saugakts. Sie wachsen also während der relativ kurzen Zeit des Saugens sehr stark an, ungefähr auf das Vierfache ihres Ursprungsvolumens. Das Wachstum geht mit feinstrukturellen Veränderungen im Zellinneren Hand in Hand, und die Zellen sondern neue Cuticulaschichten ab. In den letzten 24 Stunden des Saugakts wird das Integument dann gestreckt, wobei seine Dicke auf die Hälfte des Wertes fallen kann, der vor dem Saugen vorlag. So kann sich der Körper von Schildzecken während des Saugakts auf ein Vielfaches seines Ursprungsvolumens ausdehnen, ohne die „Haut” zu sprengen. Man vergleiche die Zeckenbeschreibung von Gaius Plinius Secundus Maior (23-79): Die Zecke zerplatzt bei Übersättigung und stirbt von ihrer Nahrung. Allerdings hält sich dieses biologisch unsinnige Gerücht hartnäckig bis in unsere Tage.
Die Beschilderung der Zecken
Die Exocuticula kann durch Einlagerung von Proteinen sklerotisieren, dh verhärten, und sich dabei braun verfärben. Insbesondere im Falle der Lederzecken sieht sie ledrig und/oder wie aus Horn gemacht aus. Dazu können noch Strukturen an der Oberfläche wie Furchen oder aufgewölbte Flächen, ⇒ Mamillen, ausgebildet werden.
Bei den Schildzecken wird auf der ⇒ Dorsalseite durch die sklerotisierte Endocuticula ein starres Rückenschild, das ⇒ Scutum beim Weibchen, das ⇒ Conscutum beim Männchen, geformt. Dieses Schild ist von Bündeln an Kanälen durchzogen, diese werden an der Oberfläche zu Poren und von uns als ⇒ Punktierung wahrgenommen. Das Schild ist nicht dehnbar, auch während und nach dem Saugakt behält es seine Form und Größe bei. Am distalen Rand des Schildes finden sich bei vielen Arten zwei zahnförmige, nach vorne gerichteten Spitzen, die ⇒ Scapulae. Form, Umriss und Größe des Schildes sind wichtige ⇒ Bestimmungsmerkmale.
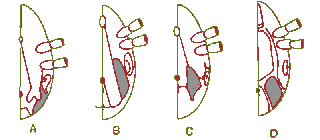
Abb. 8: Die Bauchbeschilderung von Männchen der Gattung Hyalomma (A), Boophilus (obsolete Gattung) (B), Rhipicephalus (C) und Ixodes (D).
Es existiert die Ansicht, dass die Endocuticula den Lederzecken gänzlich fehlt. Dagegen spricht aber die Ausbildung des Schildes bei den Lederzeckenlarven. In der ledrig wirkenden, meist braunen Cuticula der Lederzecken sind kleine sklerotisierte Plättchen, die Fossetten, eingebettet, an denen die Muskeln ansetzen. Auf der Oberfläche der Cuticula tragen sie außerdem kleine Schildchen, die Scutella, welche Borstenhaare (Setae) tragen. Nur die Larven der Lederzecken tragen ein unregelmäßig geformtes Schild. Lederzecken scheinen also dieses Merkmal in ihrer Stammesgeschichte verloren zu haben. Dafür spricht, dass es eine Ausnahme gibt: Die Lederzecke Nothoaspis reddelli Keirans et Clifford, 1975 ist auch als Adulttier ähnlich einer Schildzecke beschildert und besitzt keine für die Lederzecken typische weiche Cuticula am Rücken. Es ist nicht bekannt, ob diese Lederzecke - ausnahmsweise - auch eine Endocuticula besitzt.
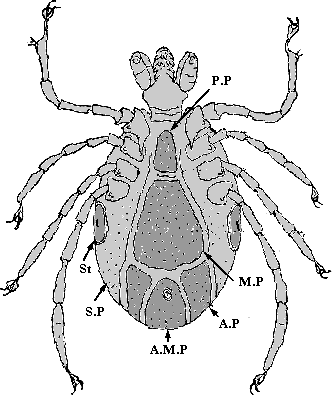
Abb. 9: Die Bauchbeschilderung eines ♂ von Ixodes ricinus. Kennungen: A.M.P: Analplatte; A.P: Paranalplatte; M.P: Mittel- oder Medianplatte; P.P: Prägenitalplatte; S.P: Epimeralplatte; St: Stigma mit Stigmenplatte.
Die Bauchseite der Schildzecken ist beschildert, während die Lederzecken dort keine nennenswerte Sklerotisierung aufweisen. Die Art der Bauchbeschilderung ist für die Gattungs- und manchmal auch für die Artbestimmung verwertbar. Besonders augenfällig ist die Bauchbeschilderung von Zecken der ⇒ prostriaten Gattung Ixodes. Im Falle dieser Gattung tragen die einzelnen Schilder eigene Namen: Der ⇒ Anus ist von der ⇒ Analplatte umgeben. Links und rechts davon liegen zwei ⇒ Adanalplatten, manchmal als Paranalplatten bezeichnet. Auf diese folgen an beiden Seiten die Lateralplatten (in Abb. 9: Epimeralplatten), welche die Beinansätze tragen, und die ⇒ Spirakularplatten, die auch Stigmenplatten oder -schilde genannt werden. Auf der Mittellinie des Bauches zwischen ⇒ Anus und ⇒ Genitalöffnung liegt die unpaarige Genitoanalplatte, auch als Mittel- oder Medianplatte bezeichnet, vor der Genitalöffnung die Prägenitalplatte. Vorne an den Seiten der Prägentialplatte liegt bei einigen wenigen Arten noch eine kleine Sternalplatte, die oft auch schon in einzelne, kleine, dreieckige oder abgerundete Plättchen aufgelöst ist. Die Nuttalliellidae stehen, auch was die Beschilderung und Sklerotisierung angeht, zwischen den Schild- und den Lederzecken.
Die Borstenhaare (Setae)
Bei den Borstenhaare (Setae) des Körpers unterscheidet man bei den Milben rund ein Dutzend verschiedener Sorten mit unterschiedlichen Funktionen. Die meisten haben Tastfunktion, einige dienen auch als Chemorezeptoren. Detailinformationen fehlen (mir) hier.
Die Körperfurchen

Abb. 10: Die Furchen auf der Dorsalseite einer Hyalomma-Art.
Sowohl die ventrale als auch die dorsale Körperoberfläche von Schildzecken ist von ⇒ Furchen durchzogen. Diese Furchen haben am Hinterleib keine erkennbare biologische Bedeutung, am Schild vergrößern sie vermutlich die Ansatzareale der Mundwerkzeugmuskulatur. Die Lage und der Verlauf dieser Furchen sind aber von hohem taxonomischem Interesse und wichtige ⇒ Bestimmungsmerkmale. Schulze [1930] schreibt, dass man vergleichend anatomisch scharf unterscheiden muss zwischen zwei Typen von Furchen:
- Den echten Furchen, Sulci genannt, die durch die Ansatzstellen der dorsiventral verlaufenden Muskulatur bedingt sind und ein Abbild der Darmdivertikeln bilden,
- und den „falschen” Furchen, Strigae genannt.
Diese Unterscheidung erscheint mir zwar zur anatomisch korrekten Bezeichnung der Furchen (zB Sulci paramediani postici) nützlich zu sein, zum Verständnis des Wesens der Tiere aber nicht viel beizutragen, und sie wurde international nicht übernommen.
III.4 Die Beine
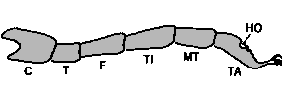
Abb. 11: Die Beinabschnitte einer Schildzecke : C: Coxa; T: Trochanter; F: Femur; TI: Tibia; MT: Metatarsus; TA: Tarsus. HO: Das Haller'sche Organ.
Milben haben mit wenigen Ausnahmen als Larven drei Beinpaare, also sechs Beine. Alle anderen Stadien besitzen vier Beinpaare, also acht Beine. Dieses Merkmal unterscheidet sie auf den ersten Blick von Insekten, die immer nur auf höchstens sechs Beinen laufen. Jedes Milbenbein besteht aus vier bis sieben, meist röhrenförmigen Abschnitten. Im Bild erkennt man am Bein einer Zecke diese Abschnitte. Es sind steife Chitinröhren, die durch dünnhäutige Verbindungen, die als Gelenke fungieren, zusammengehalten werden. Die Abschnitte werden in der Reihenfolge vom Körper nach außen wie folgt bezeichnet: ⇒ Coxa, Trochanter, Femur, Tibia, Metatarsus und Tarsus. An der Spitze jedes Beines sitzt eine Klaue, das Ambulacrum. Das Ambulacrum besteht aus einem basalen Teil, zwei Klauen in Form von Haken und einem ⇒ Pulvillus (= Empodium). Dies ist ein kissenartiges Organ, das aus feinen Härchen gebildet worden ist und den Tieren das Haften an glatten Oberflächen ermöglicht. Bei den nymphalen und adulten Argasiden (Lederzecken) fehlen die Pulvilli oder sind nur rudimentär ausgebildet. In einer Grube im ⇒ Tarsus sitzt das Haller'sche Organ, dessen Bau und Funktion weiter unten genauer beschrieben wird. Dieses Organ ist ein besonderes Merkmal des ⇒ Taxons „Milben”.
Das Coxenexterieur

Das links stehende Bild (Abb. 12) zeigt die vier Beincoxen der - ventral gesehen - rechten Körperseite eines Weibchens von Ixodes ricinus. Die Abbildung ist so orientiert, dass die vom Capitulum aus gesehene erste Coxa im Bild oben gezeigt wird und die Körpermitte des Tieres auf der linken Seite der Abbildung liegt. An den Seiten der Coxen, die zum Körperende gerichtet sind, dh den anatomischen Hinterseiten, erkennt man nach hinten gerichtete, in der Abbildung rosa eingefärbte ⇒ Dornen. Alle vier Coxen tragen einen von der Körpermitte entfernt stehenden, dh distal gelegenen Außendorn. Die erste Coxa trägt zudem auch einen ⇒ proximal stehenden Innendorn. Die Dornen dienen vermutlich der Verankerung des Blutsaugers im Haar- oder Federkleid des Wirtes, bei den Männchen auch zur Verankerung am Weibchen bei der Kopulation. Die Existenz und die Ausbildung der Innen- und Außendornen ist für die Diagnose von Ixodes-Arten ein wichtiges Merkmal.
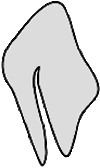 Manche Schildzecken, insbesondere die Männchen von vielen Dermacentor-Arten und beide Geschlechter in der Gattung Hyalomma, besitzen ⇒ bifide erste Coxen. Dies sind tief zweigespaltene Coxen, also Coxen mit einem tiefen Einschnitt, nicht nur mit zugespitzten Dornen. Solche Coxen dienen vermutlich der Verankerung des Tieres am Geschlechtspartner. Solch eine bifide Coxa ist rechts in Abb. 13 zu sehen.
Manche Schildzecken, insbesondere die Männchen von vielen Dermacentor-Arten und beide Geschlechter in der Gattung Hyalomma, besitzen ⇒ bifide erste Coxen. Dies sind tief zweigespaltene Coxen, also Coxen mit einem tiefen Einschnitt, nicht nur mit zugespitzten Dornen. Solche Coxen dienen vermutlich der Verankerung des Tieres am Geschlechtspartner. Solch eine bifide Coxa ist rechts in Abb. 13 zu sehen.
III.5 Die viszerale Anatomie
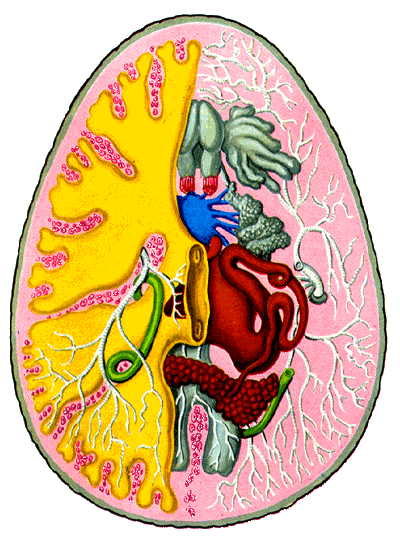 |
Abb. 14. In dieser Abbildung kann man den inneren Bauplan der heimischen Taubenzecke Argas reflexus, einer Lederzecke, studieren. cit. Grzimek [1980]. Die wichtigsten Organe und Organsysteme sind koloriert dargestellt: Graugrün und außen ist die Kutikula, die aus dem äußeren Keimblatt (Ektoderm) entstanden ist, gelb sind die aus dem inneren Keimblatt (Entoderm) entstandenen Blindsäcke des Darms, die Wohnstätten der Kommensalen, die großen, grauen, an einen Handschuh erinnernden Strukturen am Vorderende sind ein Drüsenorgan, das Gené'sche Organ, das Tracheensystem ist weiß dargestellt, eine der beiden paarigen Öffnungen ist auf der rechten Seite des Schemas dargestellt, mittelgrau sind die hinteren Blindsäcke des Enddarms, blau ist das Nervensystem, dh der Nervenknoten, karminrot die wenige Muskulatur, deren dorsiventral verlaufenden Teile im linken Bildteil querschnittig als kleine Ringe erscheinen, zinnoberrot sind die weiblichen Fortpflanzungsorgane, grün ist das wichtigste Ausscheidungsorgan, die Malpighischen Gefäße, und purpur, klein, dreieckig und fast genau zentral gelagert, aber nur auf der linken Seite sichtbar, ist das Herz. |
III.6 Die Verdauungsorgane
Der Darmtrakt
Genau wie der Darmtrakt anderer Gliedertiere besteht der von Milben aus drei Hauptteilen: Dem Vorder-, dem Mittel- und dem Enddarm. Der Vorderdarm (Stomadaeum) besteht aus der Mundöffnung, einer sklerotisierten Pharynx und dem Ösophagus. Danach folgt der Mitteldarm (Ventriculus oder Magen) mit den - falls vorhanden - daran hängenden Caeca (Blindsäcke, bläschenförmige Behälter, die die Kommensalen und Symbionten beherbergen). An den Mitteldarm schließt sich der Enddarm (Proctodaeum) an. Die Speichelgänge gehören nicht zum Verdauungstrakt, sie sind allerdings Organe, die der Verdauung der Nahrung dienen.
Der Pharynx ist elastisch und kann durch relativ starke Muskeln bewegt werden. Auf diese Weise kann der- sowohl ganz verschlossen werden, wenn die Milbe gerade keine Nahrung zu sich nimmt, als auch Saugbewegungen durchführen, mit denen flüssige Nahrung sehr effektiv in den Darm gepumpt werden kann. Feste Nahrung können Zecken nicht aufnehmen. Der Pharynx verengt sich zu einem langen Schlauch, dem Ösophagus. Dieser verläuft mitten durch das Synganglion (Gehirn) und endet im Ventriculus, dem Analogon zum Säugetiermagen. An den Ventriculus angefügt sind fünf oder, normalerweise, sieben Paare von Säckchen, die Caeca, aus denen wiederum kleinere Gänge abgehen. Diese Caeca beherbergen die Symbionten, die zur Verdauung des Blutes notwendig sind. Bei allen Zecken neigt der schmale Endabschnitt des Mitteldarms, der in den Enddarm mündet, zur Umbildung in einen lumenlosen Zellstrang und damit zum Verschluss des Darmes mit Funktionslosigkeit des After (⇒ Anus) als Kotausscheidungsorgan. Bei der Gattung Ixodes führt ein enger, aber durchgängiger Schlauch in den Enddarm. Dieser besteht aus einer Reihe hintereinander liegender Abschnitte und er endet schließlich im Anus, der im Falle der Gattung Ixodes auch (noch) genutzt wird.
Die Speicheldrüsen und -gänge
Bei den Milben besteht der Speichel absondernde Teil aus meist verzweigten Speichelgängen, die in kleine Bläschen, den Alveolen, münden. Die Zellen, die die Wände der Alveolen bilden, sondern den Speichel ab. Die Gesamtheit der Speichelgänge und Alveolen werde ich im Folgenden mit Speicheldrüsen bezeichnen. Im Mundbereich enden die Speicheldrüsen in einem Gang, der sich zwischen dem Hypostom und den ⇒ Chelizeren in den Mundraum öffnet. Prinzipiell unterscheidet man bei den Ixodida zwei Typen von Alveolen: Pyramiden-Alveolen und Sekretions-Alveolen. Die letzteren sondern den Speichel ab. Die Funktion der ersten ist nicht hundertprozentig klar, aber man vermutet, dass sie osmoregulatorisch aktiv sind. (Während des Saugakts wird das Blut angedickt: Überschüssiges Wasser und Salz wird über diese Alveolen vermutlich wieder in den Wirt abgegeben.) Der genaue Aufbau ist unterschiedlich und die Anzahl der verschiedenen Alveolen schwankt innerhalb der Ixodida. Diese Merkmale unterscheiden sich zwischen den Unterfamilien der Ixodidae. So kann man zB in der Unterfamilie Amblyominae allein vier verschiedene Typen von pyramidalen Alveolen unterscheiden. Die Lederzecken (Argasidae) haben relativ kleine Speicheldrüsen. Bei ihnen werden im Allgemeinen alle Sekrete von einem einzigen Zellentyp abgesondert. Bei den Schildzecken (Ixodidae) sind die paarigen Speichelgänge weitläufiger und rispenförmig verzweigt. Sie erstrecken sich oft bis zu den ⇒ Coxen des vierten Beinpaares. Die Speichelgänge münden in das Gnathosoma. Die Alveolen liegen in Clustern an den Seitengängen. Das Volumen der Speicheldrüsen kann während des Saugakts stark ansteigen. Der Speichel enthält außer ⇒ analgetischen und gerinnungshemmenden Stoffen auch Stoffe, die auf das Nervensystem ⇒ toxisch wirken. Diese können bei einigen Zecken-Arten zur sog. aufsteigenden Zeckenlähmung führen.
| Tab. III.1: Sekrete der Speicheldrüsen. cit. Reuben Kaufman [1989]. | |
|---|---|
| Substanz | Funktion |
| Histamin-blockierende Agentien | Nur in Homogenaten nachweisbar. Funktion unbekannt oder spekulativ. |
| Antikoagulantia | Aus verschiedenen Spezies berichtet. Es wird angenommen, dass es sich um Protein-Kohlenhydratkomplexe handelt, die die Thrombokinase hemmen. Die Antigerinnungsaktivität wurde in einigen Studien nicht gefunden. Manche Autoren behaupten, dass die Stichwunden von Zecken immer Heparin enthalten, das aus endogenen, degranulierten Mastzellen stammt. Daraus folgt, dass das Bedürfnis nach von den Zecken selbst produzierten Antigerinnungsmittel gering ist. |
| Zytolysine | Die Funktion ist, die Stichwunde zu produzieren und zu vergrößern und folglich das Saugen zu erleichtern. Im Speichel sind Esterasen und Phosphatasen gefunden worden. Obgleich eindeutige Beweise für das Vorkommen von Zytolysinen im Speichel von Lederzecken gefunden wurden, diskutieren einige Autoren ihre Anwesenheit in den Ixodiden und schlagen stattdessen vor, dass die Gewebezerstörung in der Wunde auf die Reaktion des Wirtes auf das Blutsaugen zurück zu führen ist. |
| Vasoaktive Mediatoren | Prostaglandine und einige andere Substanzen erhöhen die vaskuläre Permeabilität und erleichtern daher die Blutaufnahme. |
| Esterasen und Karbohydrat-spaltende Enzyme | Diese Enzyme erhöhen indirekt die vaskuläre Permeabilität durch Hydrolyse von Cholesterin-Estern in Mastzellen, dabei verursachen sie das Freiwerden von Histamin, 5-HT und anderen Mediatoren. |
| Paralyse-auslösende Toxine | Nachgewiesen wurden solche das Wirtstier lähmenden Toxine im Speichel von Dermacentor andersoni Stiles, 1908 und von Ixodes holocyclus Neumann, 1899. Eine für die Zecke nützliche Funktion dieser Toxine konnte bisher nicht bestimmt werden. |
Die Malpighischen Gefäße
Im Gegensatz zur ektodermalen Herkunft bei Insekten sind die „Nieren” von Zecken endodermale Bildungen. Sie heißen zwar auch Malpighischen Gefäße, sind aber keine homologen Bildungen. Es sind nur einseitig mit einem Ausgang versehene Ausstülpungen des Mitteldarmes. Im Falle der Parasitiformes handelt es sich um ein wohl entwickeltes Paar an Schläuchen, die sich im Leben lebhaft bewegen können. Die Malpighischen Gefäße dienen sowohl der Ausscheidung von kleinmolekularen Abfallstoffen als auch der Osmoregulation. Der Austransport der Stoffe erfolgt aktiv durch Transportprozesse, Filtration spielt keine Rolle. Epithelzellen transportieren Kalium- und Natrium-Ionen aus der Hämolymphe der Leibeshöhle in die Gefäße. An den Kalium-Export ist der Wasserimport gekoppelt. Aufgrund eines elektrochemischen Gradienten kann auch die schwer wasserlösliche Harnsäure in die Gefäße transportiert werden. Die Harnsäure liegt in den Gefäßen kristallin vor und spart so Lösungsmittel. Passiv werden auch Aminosäuren, Zucker, Harnstoff und andere Salzionen in das Lumen abgegeben. Der pH-Wert liegt in den Malpighischen Gefäßen bei etwa 6,8 bis 7,5. Aus dem so gebildeten isotonischen Harn werden im Enddarm aktiv Salzionen in die Hämolymphe der Leibeshöhle zurück gepumpt.
III.7 Das Blutgefäßsystem

Abb. 15: Schema des Herzens einer Schildzecke. Hellrot: Ostien; rot: Herz; gelb: Aorta. Die sechs Beinausgänge sind erkennbar, sowie rechts ein Versorgungsrohr zum Capitulum.
Viele Milben sind so klein, dass sie ein Herz und ein Blutgefäßsystem zur Versorgung ihrer Körperzellen nicht benötigen. Sie haben in diesem Fall dann auch keines ausgebildet. Über kurze Entfernungen reicht die Diffusion aus, um die Körperzellen mit Nährstoffen zu versorgen und Abfallstoffe abzutransportieren. Zecken sind aber sehr große Milben, sie benötigen ein Kreislaufsystem, das allerdings offen ist. Das Blut ist daher identisch mit der in den Organzwischenräumen befindlichen Gewebsflüssigkeit. Deshalb wird es als Hämolymphe bezeichnet. Diese enthält einige Arten kernhaltiger Zellen. Als Antriebsmotor für den Blutfluss dient ein dorso-medianer Schlauch, das Herz. Dieses wird im Laufe der Phylogenie immer weiter verkürzt. Zecken besitzen als hochentwickelte Arthropoden ein kleines, oft dreieckiges Herz mit einem Paar oder zwei Paaren von Eingangs-Öffnungen, den Ostien. Das Herz nimmt das Blut aus einem umgebenden Sinus auf und pumpt es in eine relativ lange Aorta, von welcher acht kurze Beinarterien abzweigen. Am Ende der Arterien entlassen sie das Blut wiederum in die Organzwischenräume. Der Blutfluss wird einerseits durch die Arbeit jener Muskeln erzeugt, die das Herz direkt bewegen, und andererseits derjenigen Muskeln, die Rücken und Bauch miteinander verbinden. Unten sieht man ein Schema eines Herzen einer Schildzeckenlarve. Die Strömungsrichtung des Blutes ist vom Herzen zur Aorta, also nach ⇒ kranial gerichtet, das Vorderende der Larve liegt in diesem Schema zur rechten Hand.
Die Zusammensetzung des Zeckenblutes gleicht im Prinzip der der anderen Arachniden. Im Falle der Zecken gibt es einige Untersuchungen über die verschiedenen Zelltypen, die im Blut vorkommen. Unter diesen Zellen findet man auch solche, die Abfallstoffe und Eindringlinge beseitigen können.
III.8 Das Atmungssystem
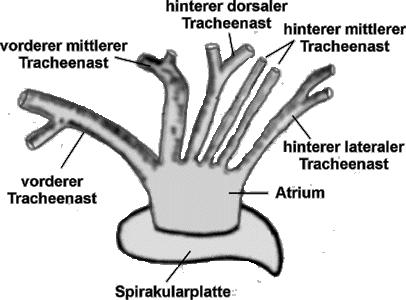
Abb. 16: Das Tracheensystem einer Schildzecke , vom Körperinneren gesehen.
Atmung bedeutet bei landlebenden Tieren die Aufnahme von Luft-Sauerstoff in den Körper und die Abgabe von Kohlendioxid. Zum Atmen bilden die höheren, landbewohnenden Arthropoden ektoblastische Einstülpungen der Cuticula aus. Diese Kanäle, Tracheen genannt, bilden eine innere Respirationsfläche. Die Ausformung der Atmungsorgane ist bei Milben recht unterschiedlich, einige Arten oder Stadien besitzen keine Tracheen. Sie nehmen den Sauerstoff vermutlich über ihr Integument auf. Die Zecken atmen als Larven auch nur über das Integument, sie besitzen als Stadien-spezifisches Merkmal keine Tracheen und daher auch keine Tracheenöffnungen. Nymphen und Adulte leiten den Luftsauerstoff mittels Diffusion durch ein einziges Paar an Tracheenröhren in den Körper. Deren äußere Öffnungen mit den Hilfsorganen heißen in der Entomologie Stigmen oder, im Falle von Spinnentieren und anderen terrestrischen Arthropoden Spiracula, auch Spirakeln. Diese liegen bei den Nuttallielliden postero-lateral zu den Coxen IV, bei den Schildzecken hinter den Coxen des vierten Beinpaares, bei den Lederzecken ventro-lateral, neben den Coxen des dritten Beinpaares. Eine oft lidförmige Struktur, die Macula, schützt den unmittelbaren Eingang der Trachee und hält diesen Eingang, das Ostium, offen. Diese Struktur liegen auf einer sklerotisierten und von Kanälen (⇒ Aeropylen, auch Poren genannt) durchzogenen Stigmen- oder, präziser, Spirakularplatte. Deren Form und deren Größe sind für die einzelnen Taxa typisch. Im Allgemeinen haben Lederzecken kleinere Spirakularplatten. Bei Schildzecken ist der Rand der Spirakularplatte sklerotisiert. Die Spirakeln regulieren einerseits den Gasaustausch und limitieren andererseits den Wasserverlust beim Atmen.
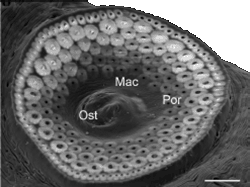
Abb. 17: Ein rasterelektronenmikroskopisches Bild des Spiraculums von Ixodes ricinus. Legende: Mac: Macula; Ost: Ostium; Por: Poren. © Suppan et al. [2013].
Die Spirakularplatten sind von zahlreichen Poren durchbrochen, welche bei den verschiedenen Zecken-Arten unterschiedliche Formen (rund, sternförmig oder andersartig) haben können und unterschiedlich angeordnet sind. Es ist nicht klar, ob diese nicht noch durch eine dünne Cuticula abgedeckt werden, deren Funktion man nicht kennt. Knapp hinter dem ⇒ proximal der Macula liegenden Atrium, einem Vorraum, verzweigen sich die Tracheen. Tracheengänge führen dann zu den Beinen und dem Synganglion, zu den Speicheldrüsen und den Geschlechtsorganen. Bei den Schildzecken führen Tracheen vom Atrium unter die Spirakularplatten. Die Wände des Atriums können durch Muskeln bewegt werden. Hierdurch wird vermutlich ein aktiver Luftaustausch bewirkt, also die Diffusion unterstützt.
III.9 Das Zentralnervensystem
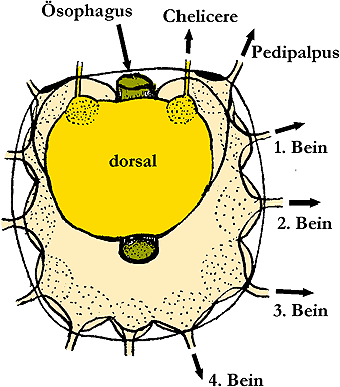
Abb. 18: Das Gehirn einer Zecke der Gattung Ornithodorus.
© Merdivenci [1969].
Das Zentralnervensystem der Tiere besteht wie bei allen Arthropoden aus dem über dem Schlund gelegenen Oberschlundganglion (= Gehirn) und dem durch Schlundkonnektive mit ihm verbundenen Bauchganglienstrang. Vom großen Oberschlundganglion gehen ein Paar Gliedmaßenganglien, die Chelizerenganglien, ab. Das dahinter liegende Ganglienpaar, jenes, das zu den Pedipalpen führt, ist nicht (mehr) homologisierbar mit jenen anderer Arthropoden , allerdings ist es sicherlich ein Teil der Unterschlundganglienkette. Diese Kette von insgesamt neun Neuromeren (2-10) ist vollständig zu einer kompakten Unterschlundmasse verschmolzen und im Prosoma konzentriert. Dadurch bildet sich ein einheitliches, großes Zentrum, von dem der ganze Körper innerviert wird. Die ⇒ Zerebralisation in der Gruppe „höher entwickelte Milben” ist deutlich erkennbar. Deren Vorteil ist offensichtlich: Kurze Wege und damit kurze Schaltzeiten. Das Zentralnervensystem der Zecken ist also kein Organ einer primitiven Gruppe, sondern ein besonders hoch entwickeltes und auf das Wesentliche reduziertes Organ eines stammesgeschichtlich alten Parasiten.
III.10 Die Sinnesorgane
Die Sinnesborsten
Die bedeutendsten Sinnesorgane der Milben sind Sinnesborsten, von denen es mehrere Typen mit unterschiedlicher Funktion gibt. Am häufigsten zu finden sind Tastborsten, andere dienen als Chemorezeptoren. Von einigen - erkennbar als Sinnesorgan genutzten - Borsten ist die Funktion ungeklärt.
Das Haller'sche Organ

Abb. 19: Ein Falschfarben-Mikrofoto des Haller'schen Organs von Ixodes ricinus.
An den vordersten Segmenten, den Tarsi, des ersten Beinpaares befindet sich ein, den Zecken und den holothyriden Milben eigenes, chemorezeptives Sinnesorgan, das Haller'sche Organ. Es wurde immer wieder behauptet, dass dieses Organ ein ⇒ autapomorphes Merkmal der Ordnung Ixodida ist, obgleich bereits in den 80-er Jahren des 20. Jahrhunderts das Vorkommen dieses Organs bei den holothyriden Milben belegt war. cit. zB Vaan der Hammen [1983]. Der Grund war das geringe Interesse der Forschergemeinde an dieser artenarmen, vorwiegend an den Rändern des Indischen Ozeans vorkommenden, vermutlichen Schwestergruppe der Ixodida.
Das Haller'sche Organ besteht aus einer mehr oder weniger flachen, manchmal an der Spitze einer Erhebung gelegenen, dann kraterförmigen Grube auf der Oberseite des ⇒ Tarsus des ersten Beinpaares und einer dahinter liegenden, in den Tarsus eingesenkten, manchmal durch ein Dach fast geschlossenen Kapsel. In beiden Strukturen stehen Sinnesborsten. Diese können manchmal so lang sein, dass sie aus der Öffnung der Kapsel hervor schauen. Diese Eigenheit kann man im Bild rechter Hand gut erkennen. Auch vor, zwischen und hinter diesen beiden Einsenkungen stehen Borstenhaare (⇒ Setae), die häufig in einer arttypischen Weise angeordnet sind. Die vordere Grube scheint eher ein Feuchtigkeitsrezeptor zu sein. Die Borstenhaare (Setae) der Kapsel dagegen dienen als Chemo- und Mechanorezeptoren. Sie stehen auf dem Grund der Kapsel und zeigen alle in Richtung der Öffnung. Das Haller'sche Organ, das alle postembryonalen Stadien, dh alle Entwicklungsstadien besitzen, ist ein komplexes sensorisches Organ, das in der olfaktorischen Wirtsfindungsaktivität der Zecke seine besondere Bedeutung hat, da es ⇒ Allomone erkennen kann. Die Rezeptoren im Haller'sche Organ ermöglichen es der Zecke, Kohlendioxid, Ammoniak, Schwefelwasserstoff und eine Vielzahl organischer Komponenten wie zB Benzaldehyd wahrzunehmen, die im Atem oder Schweiß ihrer Wirtsarten vorkommen. Vereinfacht gesagt, das Haller'sche Organ dient dazu, chemische Verbindungen- tieren, die bei der Wirtsfindung eine Rolle spielen.
Das Haller'sche Organ ist ein uraltes, von den Ursprungs-Arten stammendes Merkmal der Ixodida und der Holothyrida. Die unterschiedlichen Formen des Haller'sche Organs innerhalb der Ixodida könnten daher evolutionäre Entwicklungsschritte widerspiegeln. So nimmt man an, dass innerhalb der Lederzecken dachlose (cit.: "unroofed") Haller'sche Organe ursprüngliche, primitive Strukturen sind. cit. Hoogstraal et al. [1984]. Allerdings sind sich die Autoren in dieser Studie selbst nicht ganz sicher, ob die Dachlosigkeit nicht doch eher der Funktionalität zuzurechnen ist, schreiben sie doch ". . . the unroofed Haller's organ may be interrelated phylogenetic and biological factors."
Die Augen
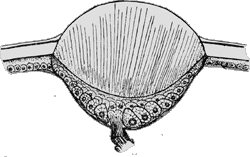
Abb. 20: Schematische Darstellung eines Zeckenauges.
© Merdivenci [1969].
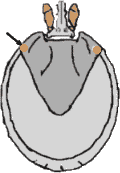
Abb. 21: Die Lage der Augen am Schildrand einer Schildzecke.
Die Augen der Arachniden bestehen aus einer Linse mit einer dahinter liegenden, halbrunden Netzhaut. Mutmaßlich waren die stammesgeschichtlich ursprünglichen Arachniden mit drei Augenpaaren ausgestattet. Im Falle der Milben sind diese sechs Augen größtenteils der sekundären Reduktion von Organen zum Opfer gefallen: Milben haben häufig gar keine Augen. Dafür finden sich bei vielen Milben Ocelli (Äuglein). So werden lichtempfindliche Stellen auf der Körperoberfläche genannt, hinter denen jedoch kein optischen Apparate stehen. Diese bilden allerdings die Umwelt nicht ab, sondern die Tiere können nur das Maß an Helligkeit erkennen. Die Ocelli sind also Tag-Nacht- und Untergrund-Höhle-Rezeptoren.
Obgleich einige mitteleuropäische Zeckengattungen (Argas, Ixodes, Haemaphysalis) gar keine erkennbaren Lichtsinnesorgane besitzen, geht man dennoch davon aus, dass Zecken aller Gattungen Licht von Dunkelheit unterscheiden können. Im Falle jener Schildzecken, die Augen besitzen, sind diese, immer nur zwei, in kleine Gruben eingesenkt und links und rechts am ⇒ lateralen Rand des ⇒ Scutums positioniert. Beäugte Lederzecken hingegen haben die Augen an der ventro-lateralen Körperoberfläche, lateral zu den Beinen. Vor allem die Pirsch-jagenden „Laufzecken” scheinen ihre Wirte mittels ihrer Augen wahrzunehmen.
III.11 Das Geschlecht
Die Geschlechtsbestimmung
Die Geschlechtsbestimmung erfolgt bei den Milben durch recht unterschiedliche Methoden, bei den Zecken wird das Geschlecht durch Geschlechtschromosomen (Gonosomen) festgelegt. Diese sind meist deutlich größer als die anderen Chromosomen, die Autosomen, und an ihrer Größe leicht zu erkennen. Die Weise der Geschlechtsdetermination ist unterschiedlich: Viele Schildzecken-Arten und die Lederzecken besitzen X und Y-Chromosomen. Weibchen sind durch XX determiniert und daher homozygot, das Männchen durch XY (XX:XY-System). Nach anderen Autoren besitzen Schildzecken nur X-Gonosomen, Weibchen haben zwei davon, also XX, Männchen hingegen nur eines (XX:X0-System). Die Art Haemaphysalis sulcata ist detaillierter studiert worden: Der diploide Chromosomensatz des Weibchens beträgt (2n =) 22 und die Geschlechtsbestimmung erfolgt durch das XX:XO-System. cit. Valero et al. [1997]. Dies führt dazu, dass Männchen ein Chromosom weniger besitzen als Weibchen; in diesem Fall also 2n = 21. cit. Oliver [1964]. Wieder andere Schildzecken besitzen vielleicht mehrere Geschlechtschromosomen (zB Typ X1X1X2X2:X1X2Y). Die Realisation dieser Möglichkeiten schwankt nicht nur innerhalb der Familien und Gattungen, möglicherweise können sogar bei verschiedenen Populationen einer einzigen Art Unterschiede in der Geschlechtsbestimmung auftreten. Im Falle der Verwirklichung dieser Annahme ist allerdings der Genfluss innerhalb der „Art” unterbrochen und damit definitionsgemäß die Existenz mehrere Arten bewiesen, die Kryptoarten sind.
Die sekundären Geschlechtsmerkmale
Der Unterschied zwischen den Geschlechtern in den sekundären Geschlechtsmerkmalen ist bei den Lederzecken (Argasidae) marginal. Bei den Schildzecken (Ixodidae) sind die Männchen im Allgemeinen etwas kleiner als die Weibchen. Ihr Rücken ist vollständig vom Scutum bedeckt, während es bei den Weibchen nur einen kleinen Teil des vorderen Rückens verdeckt. Bei etlichen Arten ist bei den Männchen das ⇒ Hypostom verkümmert oder ungewöhnlich pilzförmig geformt. Auch besitzen nur die Weibchen der Schildzecken ⇒ Areae porosae.
Die weiblichen Geschlechtsorgane
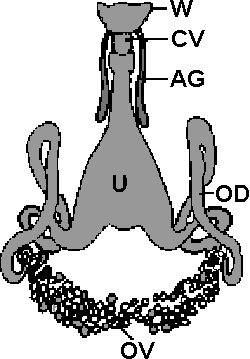
Abb. 22: Der Geschlechtstrakt eines Weibchens der Lederzecke Argas persicus. Abbildungslegende: VV: Vestibulum vaginae; AG: akzessorische Drüsen; CV: Uteruszugang; U: Uterus; OD: Ovidukt; OV: Ovarien.
Im Geschlechtstrakt einer weiblichen Lederzecke folgt hinter dem Vaginaleingang das Vestibulum vaginae eine Art Vorraum, in das die akzessorischen Drüsen münden. Dahinter folgen dann der Uteruszugang und der Uterus. Von diesem gehen zwei Ovidukte oder Eileiter ab, die schließlich in den Ovarien oder Eierstöcken enden. Bei den Schildzecken sind die Ovarien ebenso wie die Ovidukte asymmetrisch angelegt und stark gewunden.
Wenn das Weibchen Blut gesogen hat, setzen sich Oozyten oder Eimutterzellen an den Wände der Ovarien fest und fangen an auszureifen. Während dieser Zeit verlängert sich das Ovarium und fängt an, sich zu falten und zu winden. Die reifen Oozyten, das sind die unbefruchteten Eier, lösen sich von den Wänden der Ovarien und sammeln sich in der Flüssigkeit.
Die Anzahl der Eier ist sehr unterschiedlich. Schildzecken, die nur einmal in ihrem Leben Eier legen, können sehr viele produzieren; Ixodes ricinus (Linnaeus, 1758) legt mehrere tausend Eier, angeblich bis zu 10 000. Lederzecken dagegen legen deutlich weniger, 50 bis einige hundert, dafür aber mehrfach.
Das Gené'sche Organ
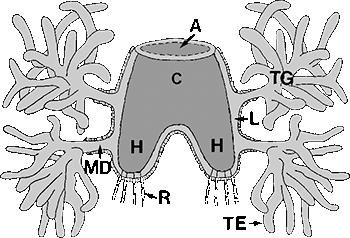
Abb. 23: Das Gené'sche Organ von Dermacentor reticulatus. Abbildungslegende: A: Öffnung; C: Corpus = kutikulärer Sack ; H: Hörner; L: Lumen zwischen dem Epithelial- und dem Cuticularsack; MD: Hauptausführgang der Drüsen; TE: tubuläre Endstücke; TG: tubuläre Drüsen; R: Retraktormuskeln.
© Schöl et al. [2001].
In wahrscheinlich allen adulten, weiblichen Zecken findet sich eine Drüse, das Gené'sches Organ, mit Hilfe derer die Zecke ihre Eier mit einer Wachshülle versieht. Diese Drüse ist einzigartig in der ganzen Tierwelt, es handelt sich also um ein ⇒ autapomorphes Merkmal der Zecken. Das Gené'sche Organ besteht aus einem Flaschenboden-förmigen Innenraum, in den zwei (Lederzecken?) oder vier Ausführgänge der Drüsen münden. Im Innenraum befindet sich ein zylindrischer „Corpus” aus nicht näher definiertem Material, der ⇒ proximal zwei, nach anderen Meinungen vier, Hörner ausbildet. Distal liegt die ovale Öffnung nach außen. Diese Öffnung des Gené'schen Organs liegt in einer Spalte hinter dem postero-dorsalen Rand der
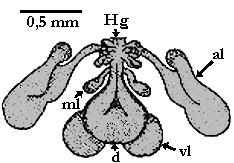
Abb. 24: Das Gené'sche Organ von Nuttallielliden. Abbildungslegende: al: antero-laterales Drüsen-Taschenpaar (DTp); ml: mittig-laterales DTp; vl: ventro-laterales DTp; d: Dorsaltasche; Hg: Hauptgang.
© El Shoura et al. [1984].
Das Gené'sche Organ der Nuttalliellidae ist in seinem Bau einzigartig innerhalb der Zecken. Es liegt unterhalb der Position am Rücken, an der beim Männchen das Pseudoscutum liegt, und es besteht aus sieben Drüsen-Taschen: Eine einzelne, die dorsale, und drei in Paaren angeordnete: Ein antero-laterales, ein mittig-laterales und ein ventro-laterales Paar. Diese münden in einen Hauptgang, der nach außen führt. Er endet in einem transversalen Schlitz in der dorsalen Körperwand. Dieser Schlitz liegt dort, wo beim Männchen der Vorderrand des Pseudoscutums und der postero-dorsale Rand der ⇒ Basis capituli liegen. Vor der Außenöffnung ist der Hauptgang noch von einigen kleinen Bläschen umgeben, die sich durch enge Verbindungen in ihn hinein öffnen. cit. El Shoura et al. [1984]. Es ist nicht bekannt, ob dieses Organ in gleicher Weise wie unten beschrieben seine Aufgabe im Rahmen des Eierlegens erfüllen kann oder aber eine einfacher konstruierte Vorläuferstruktur mit gleichen oder ähnlichen Funktionen ist. Ich meine, dass eine Reduktion von sieben Drüsensäcken zu vier ähnlich aussehenden, paarig angelegten tubulären Drüsen kaum vorstellbar ist. Mit dieser Ansicht ergibt sich zwingend, dass das Gené'sche Organ der Nuttalliellidae keine Vorläuferstruktur aller Gené'schen Organe ist, sondern eine eigenständige Entwicklung.
Der Prozess der Eiablage ist eine stereotype Sequenz von mehreren, zeitlich exakt koordinierten, aufeinander folgenden und ineinander greifenden Vorgängen. Zum Eierlegen krümmt sich das Weibchen nach unten und hinten und bringt sein ⇒ Capitulum unter den Bauch in Richtung der ⇒ Genitalöffnung. Die Eiablage wird durch dieses Absenken des Capitulums eingeleitet, gleichzeitig werden die Pedipalpen gespreizt und bis zur ventralen Körperwand abgesenkt. So wird die Geschlechtsöffnung beidseitig umfasst. Unmittelbar daran schließt sich mehrfach ein Aus- und Einstülpen des Corpus des Gené'schen Organs an, das schließlich in einer maximalen Entfaltung des kutikulären Sacks (= Corpus) endet. Das Vestibulum vaginae wölbt sich nun röhrenförmig vor, wird zu einem Ovipositor und übergibt ein Ei nach einem kurzen, breitflächigen Kontakt mit dem kutikulären Sack an dessen beiden Hörner. Das austretende Ei wird etwa zwei Minuten lang von den Pedipalpen (?) um die Hörner des Gené'sche Organs gedreht und so mit einer Wachshülle überzogen. Danach wird der Ovipositor wieder zurückgezogen, die Pedipalpen werden geschlossen und der kutikuläre Sack retrahiert. In der folgenden Endphase des Eiablageprozesses befördert die Zecke das Ei mittels einer kräftigen Aufwärtsbewegung des Capitulums auf ihren Rücken oder - möglicherweise artabhängig - vor sich auf den Boden. Das Capitulum kehrt auf seinen ursprünglichen Platz zurück. Eier, denen die Wachsschicht fehlt, haben nur minimale Überlebenschancen. Sie trocknen aus und schrumpfen - es wird allerdings auch behauptet, dass diese Wachsschicht ein starkes, räuberische-Milben-tötendes Gift enthält. Verändert cit. Schöl et al. [2001].
Die männlichen Geschlechtsorgane

Abb. 25: Der Geschlechtstrakt eines Männchen von Dercamentor andersoni. Abbildungslegende: TE: Testes; VD: Vas deferens; AG: akzessorische Drüsen; EjD: Ejakulationsgang.
Der männliche Geschlechtstrakt besteht aus den folgenden Teilen: Den Testes oder Hoden, einem Vas deferens oder Samenleiter, den akzessorische Drüsen und einem Ejakulationsgang. Männliche Zecken besitzen zwei paarig angeordnete Hoden, welche in der Gruppe der Lederzecken an ihrem Ende durch eine kleine Brücke verbunden sind. An ihrem unteren Ende verengen sich die Hoden zu den Samenleitern. In der ⇒ proximalen Spitze der Hoden werden die Spermatogone gebildet, die dann die Testes hinabwandern, sich dabei im Inneren der Hoden teilen und schließlich Zysten bilden, in deren Inneren die Samenzellen gebildet werden. Diese Zysten wandern in die Vasa deferentia ein, welche als Samengänge und -behälter dienen. Dort löst sich die Membran, die die Zyste zusammenhält, auf und eine sekundäre Zyste wird gebildet, in der schließlich die Samenzellen endgültig ausreifen. Während der Paarung wandern diese sekundären Zysten in den Ejakulationsgang, mischen sich dort mit der Flüssigkeit aus den akzessorischen Drüsen und werden in eine ⇒ Spermatophore eingeschlossen. Die Spermatophore wird abgesetzt und härtet beim Luftkontakt aus.